

Centriole duplication: analogue control in a digital age
- PMID: 21067522
- PMCID: PMC3051170
- DOI: 10.1042/CBI20100612
Centriole duplication: analogue control in a digital age
Abstract
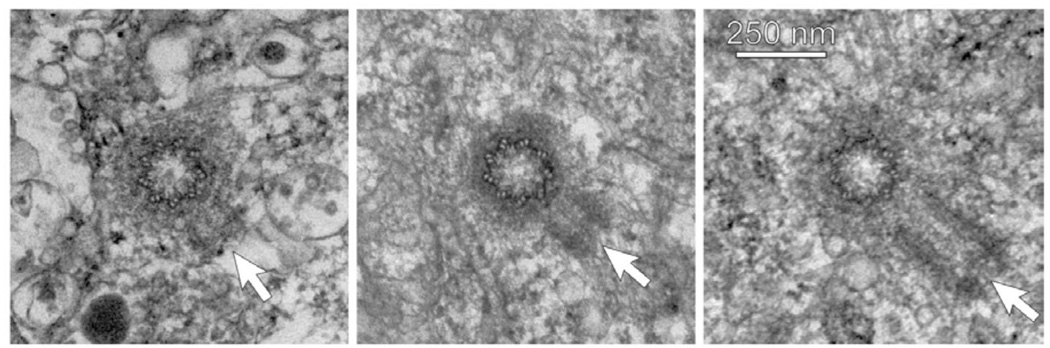
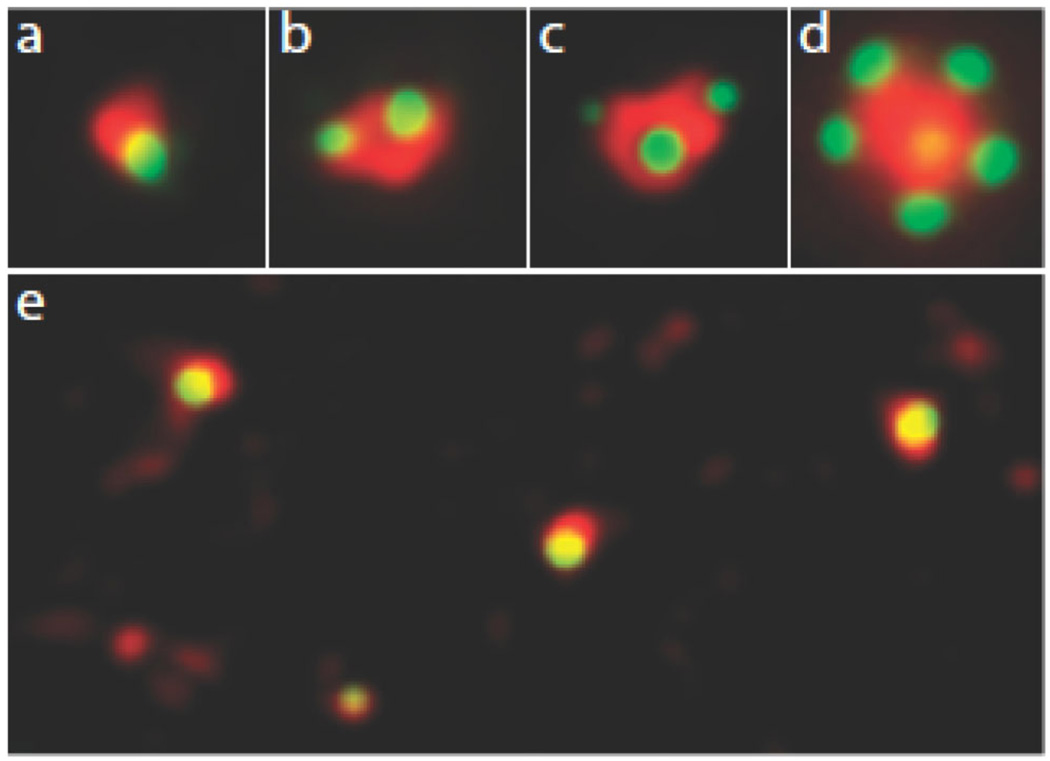
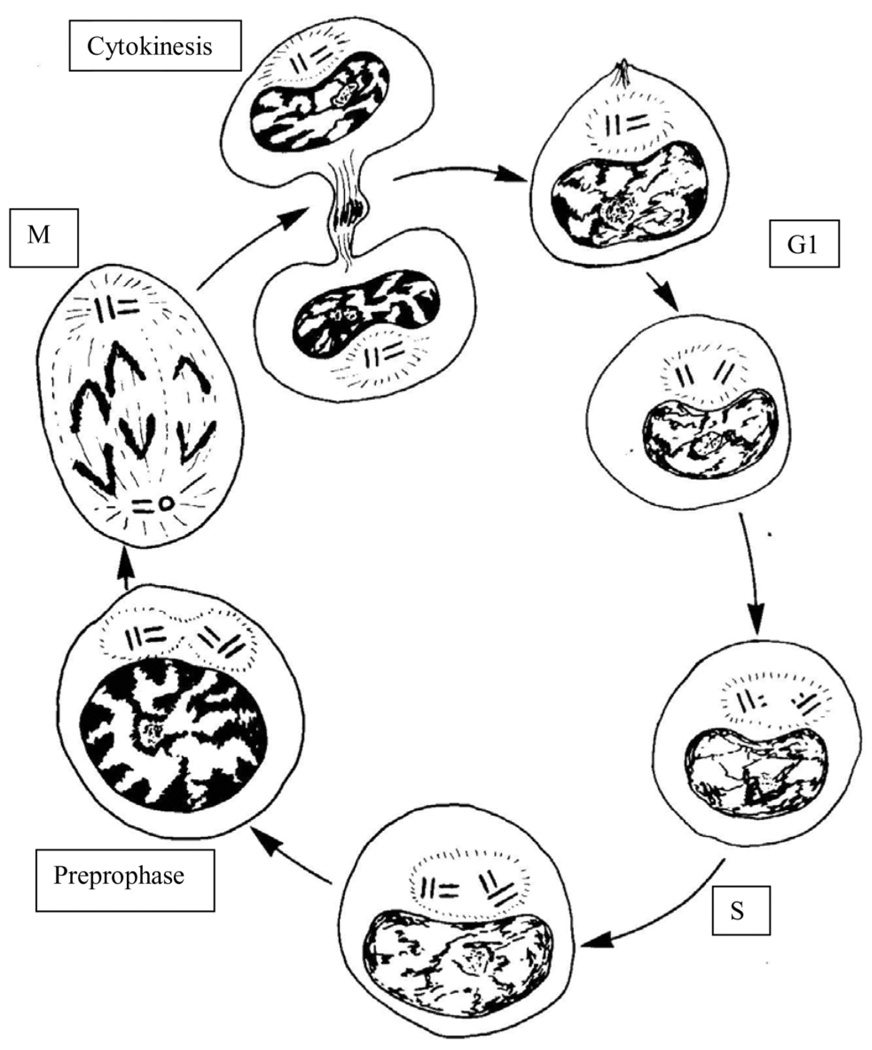
In preparation for mitosis, the centrosome doubles once and only once to provide the two poles of the mitotic spindle. The presence of more than two centrosomes increases the chances that mitosis will be multipolar, and chromosomes will be distributed unequally. Since the number of mother-daughter centriole pairs determines the number of centrosomes, it is important that only one daughter centriole is assembled at, but slightly separated from, the proximal end of each mother centriole. This numerical and spatial specificity has led to the belief that a 'template' on the mother centriole provides a unique site for procentriole assembly. We review observations that are leading to the demise of this intuitively attractive idea. In its place, we are left with the notion that pericentriolar material at the wall of the mother centriole provides a local environment that promotes the assembly of a macromolecular complex that seeds the daughter centriole. Even though the system normally behaves in a digital fashion to go from zero to just one daughter centriole per mother, this behaviour appears to be based in the precise analogue control of multiple proteins, their activities, and the structure provided by the mother centriole.
Figures
Similar articles
-
Centriole inheritance.Prion. 2008 Jan-Mar;2(1):9-16. doi: 10.4161/pri.2.1.5064. Epub 2008 Jan 12. Prion. 2008. PMID: 19164929 Free PMC article. Review.
-
One to only two: a short history of the centrosome and its duplication.Philos Trans R Soc Lond B Biol Sci. 2014 Sep 5;369(1650):20130455. doi: 10.1098/rstb.2013.0455. Philos Trans R Soc Lond B Biol Sci. 2014. PMID: 25047609 Free PMC article. Review.
-
ASQ2 encodes a TBCC-like protein required for mother-daughter centriole linkage and mitotic spindle orientation.Curr Biol. 2009 Jul 28;19(14):1238-43. doi: 10.1016/j.cub.2009.05.071. Curr Biol. 2009. PMID: 19631545 Free PMC article.
-
The centrosome cycle: Centriole biogenesis, duplication and inherent asymmetries.Nat Cell Biol. 2011 Oct 3;13(10):1154-60. doi: 10.1038/ncb2345. Nat Cell Biol. 2011. PMID: 21968988 Free PMC article. Review.
-
PCNT is critical for the association and conversion of centrioles to centrosomes during mitosis.J Cell Sci. 2019 Mar 26;132(6):jcs225789. doi: 10.1242/jcs.225789. J Cell Sci. 2019. PMID: 30814333
Cited by
-
Centrioles and Ciliary Structures during Male Gametogenesis in Hexapoda: Discovery of New Models.Cells. 2020 Mar 18;9(3):744. doi: 10.3390/cells9030744. Cells. 2020. PMID: 32197383 Free PMC article. Review.
-
Let's huddle to prevent a muddle: centrosome declustering as an attractive anticancer strategy.Cell Death Differ. 2012 Aug;19(8):1255-67. doi: 10.1038/cdd.2012.61. Epub 2012 Jun 1. Cell Death Differ. 2012. PMID: 22653338 Free PMC article. Review.
-
A cell cycle role for the epigenetic factor CTCF-L/BORIS.PLoS One. 2012;7(6):e39371. doi: 10.1371/journal.pone.0039371. Epub 2012 Jun 19. PLoS One. 2012. PMID: 22724006 Free PMC article.
-
Exploring the evolutionary history of centrosomes.Philos Trans R Soc Lond B Biol Sci. 2014 Sep 5;369(1650):20130453. doi: 10.1098/rstb.2013.0453. Philos Trans R Soc Lond B Biol Sci. 2014. PMID: 25047607 Free PMC article. Review.
-
Centrosome dysfunction contributes to chromosome instability, chromoanagenesis, and genome reprograming in cancer.Front Oncol. 2013 Nov 12;3:277. doi: 10.3389/fonc.2013.00277. eCollection 2013. Front Oncol. 2013. PMID: 24282781 Free PMC article. Review.
References
-
- Alvey PL. An investigation of the centriole cycle using 3T3 and CHO cells. J Cell Sci. 1985;78:147–162. - PubMed
-
- Andersen JS, Wilkinson CJ, Mayor T, Mortensen P, Nigg EA, Mann M. Proteomic characterization of the human centrosome by protein correlation profiling. Nature. 2003;426:570–574. - PubMed
-
- Beisson J, Wright M. Basal body/centriole assembly and continuity. Curr Opin Cell Biol. 2003;15:96–104. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
