

Parasitoidism, not sociality, is associated with the evolution of elaborate mushroom bodies in the brains of hymenopteran insects
- PMID: 21068044
- PMCID: PMC3049053
- DOI: 10.1098/rspb.2010.2161
Parasitoidism, not sociality, is associated with the evolution of elaborate mushroom bodies in the brains of hymenopteran insects
Abstract
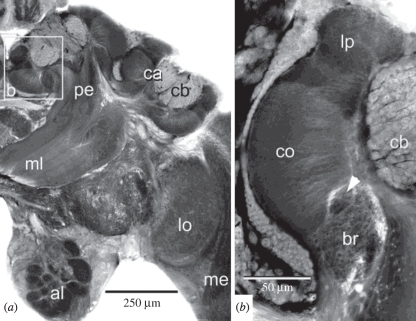
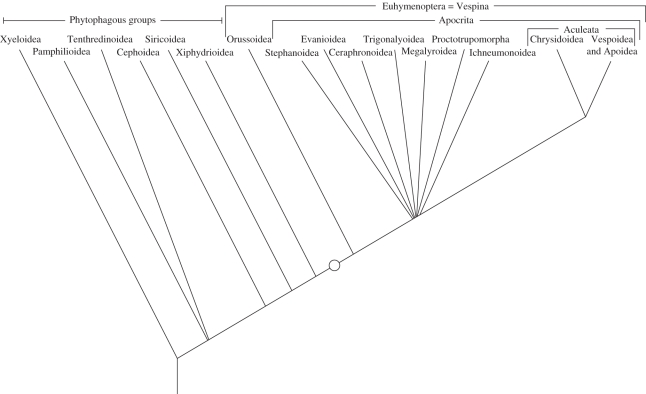
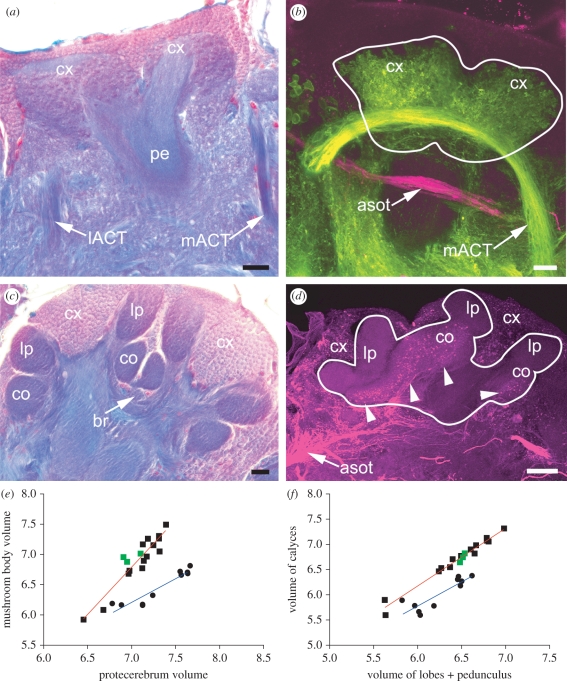
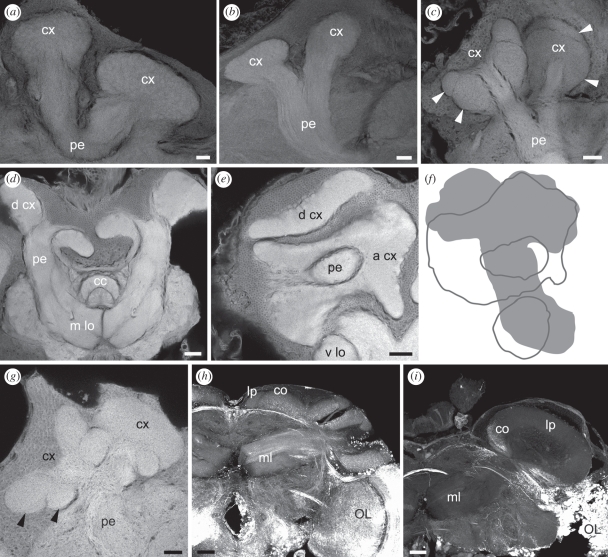
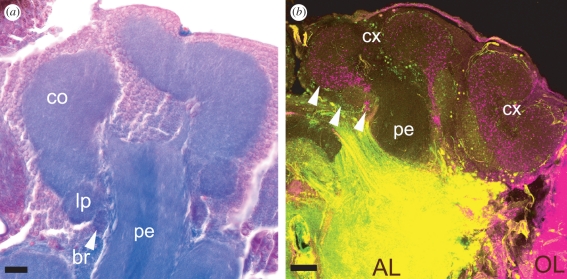
The social brain hypothesis posits that the cognitive demands of social behaviour have driven evolutionary expansions in brain size in some vertebrate lineages. In insects, higher brain centres called mushroom bodies are enlarged and morphologically elaborate (having doubled, invaginated and subcompartmentalized calyces that receive visual input) in social species such as the ants, bees and wasps of the aculeate Hymenoptera, suggesting that the social brain hypothesis may also apply to invertebrate animals. In a quantitative and qualitative survey of mushroom body morphology across the Hymenoptera, we demonstrate that large, elaborate mushroom bodies arose concurrent with the acquisition of a parasitoid mode of life at the base of the Euhymenopteran (Orussioidea + Apocrita) lineage, approximately 90 Myr before the evolution of sociality in the Aculeata. Thus, sociality could not have driven mushroom body elaboration in the Hymenoptera. Rather, we propose that the cognitive demands of host-finding behaviour in parasitoids, particularly the capacity for associative and spatial learning, drove the acquisition of this evolutionarily novel mushroom body architecture. These neurobehavioural modifications may have served as pre-adaptations for central place foraging, a spatial learning-intensive behaviour that is widespread across the Aculeata and may have contributed to the multiple acquisitions of sociality in this taxon.
Figures
References
-
- McGuire S. E., Le P. T., Davis R. L. 2001. The role of Drosophila mushroom body signaling in olfactory memory. Science 293, 1330–1333 10.1126/science.1062622 (doi:10.1126/science.1062622) - DOI - PubMed
-
- Perez-Orive J., Mazor O., Turner G. C., Cassenaer S., Wilson R. I., Laurent G. 2002. Oscillations and sparsening of odor representation in the mushroom body. Science 297, 359–365 10.1126/science.1070502 (doi:10.1126/science.1070502) - DOI - PubMed
-
- Blum A. L., Li W., Cressy M., Dubnau J. 2009. Short- and long-term memory in Drosophila require cAMP signaling in distinct cell types. Curr. Biol. 19, 1341–1350 10.1016/j.cub.2009.07.016 (doi:10.1016/j.cub.2009.07.016) - DOI - PMC - PubMed
-
- Mizunami M., Weibrecht J. M., Strausfeld N. J. 1993. A new role for the insect mushroom bodies: place memory and motor control. In Biological neural networks in invertebrate neuroethology and robotics (eds Beer R. D., Ritzman R. E., McKenna T.), pp. 199–225 New York, NY: Academic Press
-
- Mizunami M., Weibrecht J. M., Strausfeld N. J. 1998. Mushroom bodies of the cockroach: their participation in place memory. J. Comp. Neurol. 402, 520–537 10.1002/(SICI)1096-9861(19981228)402:4<520::AID-CNE6>3.0.CO;2-K (doi:10.1002/(SICI)1096-9861(19981228)402:4<520::AID-CNE6>3.0.CO;2-K) - DOI - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources