

dFMRP and Caprin, translational regulators of synaptic plasticity, control the cell cycle at the Drosophila mid-blastula transition
- PMID: 21068064
- PMCID: PMC2990211
- DOI: 10.1242/dev.055046
dFMRP and Caprin, translational regulators of synaptic plasticity, control the cell cycle at the Drosophila mid-blastula transition
Abstract
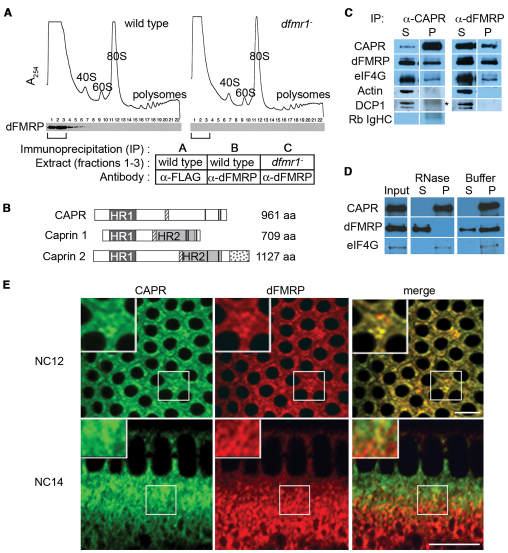
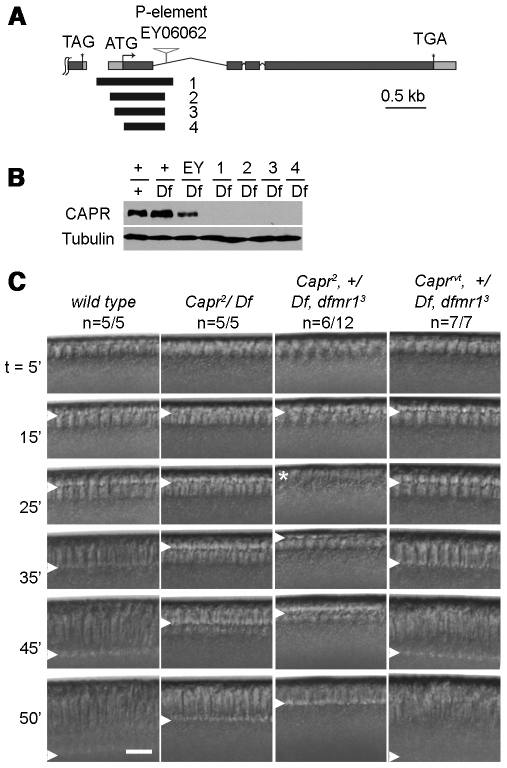
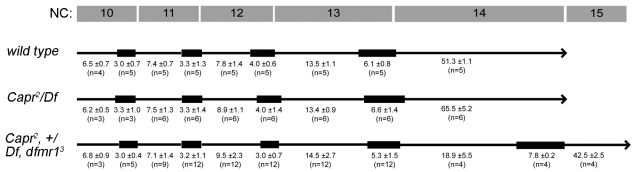
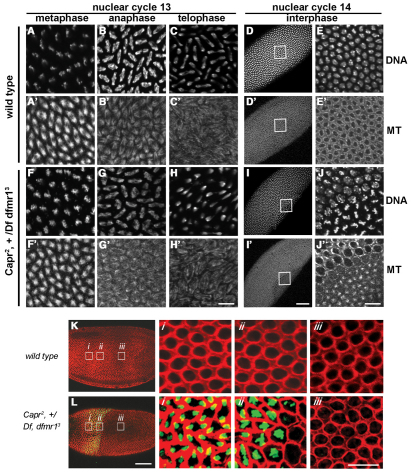
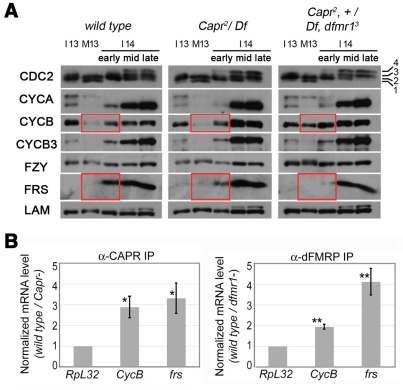
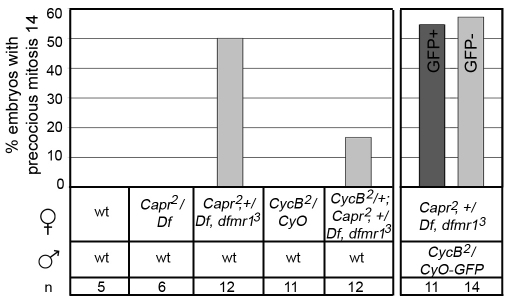
The molecular mechanisms driving the conserved metazoan developmental shift referred to as the mid-blastula transition (MBT) remain mysterious. Typically, cleavage divisions give way to longer asynchronous cell cycles with the acquisition of a gap phase. In Drosophila, rapid synchronous nuclear divisions must pause at the MBT to allow the formation of a cellular blastoderm through a special form of cytokinesis termed cellularization. Drosophila Fragile X mental retardation protein (dFMRP; FMR1), a transcript-specific translational regulator, is required for cellularization. The role of FMRP has been most extensively studied in the nervous system because the loss of FMRP activity in neurons causes the misexpression of specific mRNAs required for synaptic plasticity, resulting in mental retardation and autism in humans. Here, we show that in the early embryo dFMRP associates specifically with Caprin, another transcript-specific translational regulator implicated in synaptic plasticity, and with eIF4G, a key regulator of translational initiation. dFMRP and Caprin collaborate to control the cell cycle at the MBT by directly mediating the normal repression of maternal Cyclin B mRNA and the activation of zygotic frühstart mRNA. These findings identify two new targets of dFMRP regulation and implicate conserved translational regulatory mechanisms in processes as diverse as learning, memory and early embryonic development.
Figures
References
-
- Aerbajinai W., Lee Y. T., Wojda U., Barr V. A., Miller J. L. (2004). Cloning and characterization of a gene expressed during terminal differentiation that encodes a novel inhibitor of growth. J. Biol. Chem. 279, 1916-1921 - PubMed
-
- Anderson P., Kedersha N. (2009). RNA granules: post-transcriptional and epigenetic modulators of gene expression. Nat. Rev. Mol. Cell Biol. 10, 430-436 - PubMed
-
- Arbeitman M. N., Furlong E. E., Imam F., Johnson E., Null B. H., Baker B. S., Krasnow M. A., Scott M. P., Davis R. W., White K. P. (2002). Gene expression during the life cycle of Drosophila melanogaster. Science 297, 2270-2275 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
