

Src-family and Syk kinases in activating and inhibitory pathways in innate immune cells: signaling cross talk
- PMID: 21068150
- PMCID: PMC3039931
- DOI: 10.1101/cshperspect.a002352
Src-family and Syk kinases in activating and inhibitory pathways in innate immune cells: signaling cross talk
Abstract
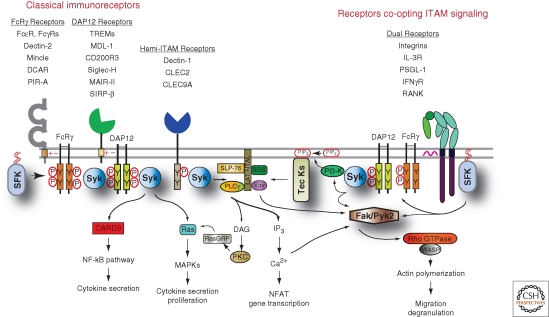
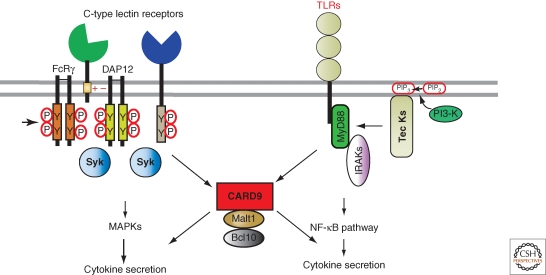
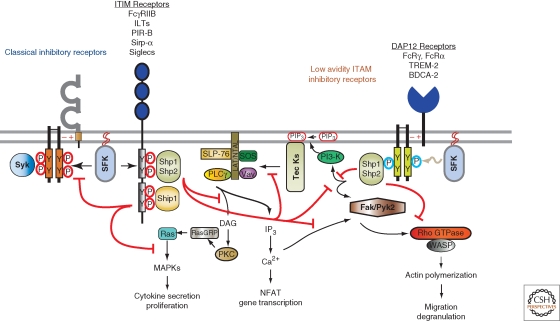
The response of innate immune cells to growth factors, immune complexes, extracellular matrix proteins, cytokines, pathogens, cellular damage, and many other stimuli is regulated by a complex net of intracellular signal transduction pathways. The majority of these pathways are either initiated or modulated by Src-family or Syk tyrosine kinases present in innate cells. The Src-family kinases modulate the broadest range of signaling responses, including regulating immunoreceptors, C-type lectins, integrins, G-protein-coupled receptors, and many others. Src-family kinases also modulate the activity of other kinases, including the Tec-family members as well as FAK and Pyk2. Syk kinase is required for initiation of signaling involving receptors that utilize immunoreceptor tyrosine activation (ITAM) domains. This article reviews the major activating and inhibitory signaling pathways regulated by these cytoplasmic tyrosine kinases, illuminating the many examples of signaling cross talk between pathways.
Figures
Similar articles
-
Src and Syk kinases: key regulators of phagocytic cell activation.Trends Immunol. 2005 Apr;26(4):208-14. doi: 10.1016/j.it.2005.02.002. Trends Immunol. 2005. PMID: 15797511 Review.
-
Protein tyrosine kinases Syk and ZAP-70 display distinct requirements for Src family kinases in immune response receptor signal transduction.J Immunol. 1997 Feb 15;158(4):1650-9. J Immunol. 1997. PMID: 9029101
-
G6b-B inhibits constitutive and agonist-induced signaling by glycoprotein VI and CLEC-2.J Biol Chem. 2008 Dec 19;283(51):35419-27. doi: 10.1074/jbc.M806895200. Epub 2008 Oct 27. J Biol Chem. 2008. PMID: 18955485 Free PMC article.
-
The Src, Syk, and Tec family kinases: distinct types of molecular switches.Cell Signal. 2010 Aug;22(8):1175-84. doi: 10.1016/j.cellsig.2010.03.001. Epub 2010 Mar 4. Cell Signal. 2010. PMID: 20206686 Review.
-
Functional interaction of protein kinase Calpha with the tyrosine kinases Syk and Src in human platelets.J Biol Chem. 2005 Feb 25;280(8):7194-205. doi: 10.1074/jbc.M409212200. Epub 2004 Dec 6. J Biol Chem. 2005. PMID: 15583006
Cited by
-
The uric acid crystal receptor Clec12A potentiates type I interferon responses.Proc Natl Acad Sci U S A. 2019 Sep 10;116(37):18544-18549. doi: 10.1073/pnas.1821351116. Epub 2019 Aug 26. Proc Natl Acad Sci U S A. 2019. PMID: 31451663 Free PMC article.
-
De novo identification of maximally deregulated subnetworks based on multi-omics data with DeRegNet.BMC Bioinformatics. 2022 Apr 19;23(1):139. doi: 10.1186/s12859-022-04670-6. BMC Bioinformatics. 2022. PMID: 35439941 Free PMC article.
-
B cell-specific loss of Lyn kinase leads to autoimmunity.J Immunol. 2014 Feb 1;192(3):919-28. doi: 10.4049/jimmunol.1301979. Epub 2013 Dec 27. J Immunol. 2014. PMID: 24376269 Free PMC article.
-
SYK inhibition blocks proliferation and migration of glioma cells and modifies the tumor microenvironment.Neuro Oncol. 2018 Apr 9;20(5):621-631. doi: 10.1093/neuonc/noy008. Neuro Oncol. 2018. PMID: 29401256 Free PMC article.
-
Reporter cell assay for human CD33 validated by specific antibodies and human iPSC-derived microglia.Sci Rep. 2021 Jun 29;11(1):13462. doi: 10.1038/s41598-021-92434-2. Sci Rep. 2021. PMID: 34188106 Free PMC article.
References
-
- Abdelbaqi M, Chidlow JH, Matthews KM, Pavlick KP, Barlow SC, Linscott AJ, Grisham MB, Fowler MR, Kevil CG 2006. Regulation of dextran sodium sulfate induced colitis by leukocyte beta 2 integrins. Lab Invest 86: 380–390 - PubMed
-
- Amoras AL, Kanegane H, Miyawaki T, Vilela MM 2003. Defective Fc-, CR1- and CR3-mediated monocyte phagocytosis and chemotaxis in common variable immunodeficiency and X-linked agammaglobulinemia patients. J Investig Allergol Clin Immunol 13: 181–188 - PubMed
-
- Arefieva TI, Kukhtina NB, Antonova OA, Krasnikova TL 2005. MCP-1-stimulated chemotaxis of monocytic and endothelial cells is dependent on activation of different signaling cascades. Cytokine 31: 439–446 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous