

Integration of clearance mechanisms: the proteasome and autophagy
- PMID: 21068151
- PMCID: PMC2982176
- DOI: 10.1101/cshperspect.a006734
Integration of clearance mechanisms: the proteasome and autophagy
Abstract
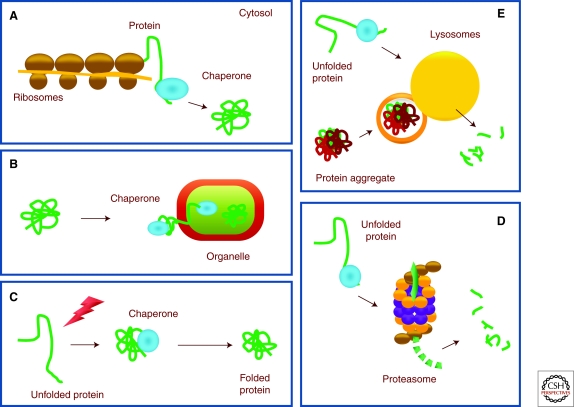
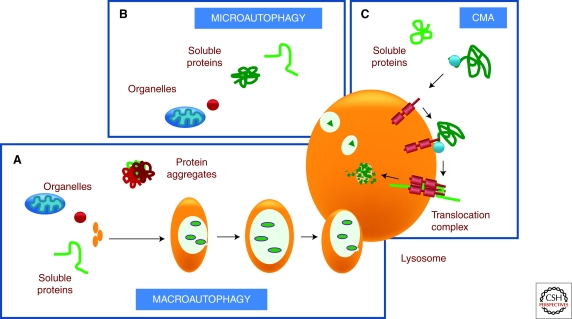
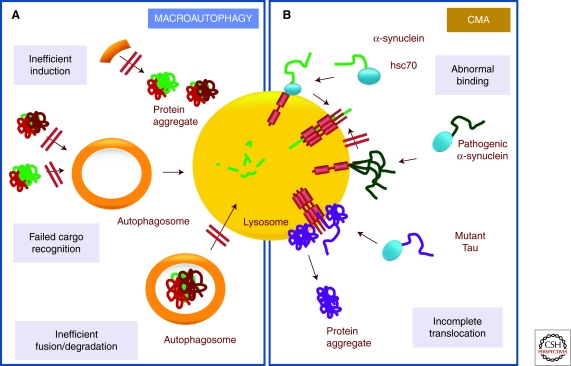
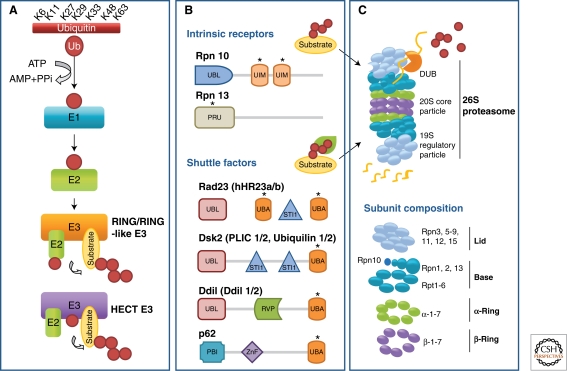
Cells maintain a healthy proteome through continuous evaluation of the quality of each of their proteins. Quality control requires the coordinated action of chaperones and proteolytic systems. Chaperones identify abnormal or unstable conformations in proteins and often assist them to regain stability. However, if repair is not possible, the aberrant protein is eliminated from the cellular cytosol to prevent undesired interactions with other proteins or its organization into toxic multimeric complexes. Autophagy and the ubiquitin/proteasome system mediate the complete degradation of abnormal protein products. In this article, we describe each of these proteolytic systems and their contribution to cellular quality control. We also comment on the cellular consequences resulting from the dysfunction of these systems in common human protein conformational disorders and provide an overview on current therapeutic interventions based on the modulation of the proteolytic systems.
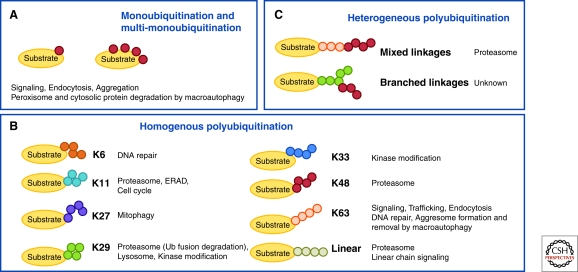
Figures
References
-
- Adams J 2004. The development of proteasome inhibitors as anticancer drugs. Cancer Cell 5: 417–421 - PubMed
-
- Ahlberg J, Glaumann H 1985. Uptake–microautophagy–and degradation of exogenous proteins by isolated rat liver lysosomes. Effects of pH, ATP, and inhibitors of proteolysis. Exp Mol Pathol 42: 78–88 - PubMed
-
- Alves G, Forsaa EB, Pedersen KF, Dreetz Gjerstad M, Larsen JP 2008. Epidemiology of Parkinson's disease. J Neurol 255: 18–32 - PubMed
-
- Baboshina OV, Haas AL 1996. Novel multiubiquitin chain linkages catalyzed by the conjugating enzymes E2EPF and RAD6 are recognized by 26S proteasome subunit 5. J Biol Chem 271: 2823–2831 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources