

Native and recombinant Slc26a3 (downregulated in adenoma, Dra) do not exhibit properties of 2Cl-/1HCO3- exchange
- PMID: 21068358
- PMCID: PMC3043631
- DOI: 10.1152/ajpcell.00366.2010
Native and recombinant Slc26a3 (downregulated in adenoma, Dra) do not exhibit properties of 2Cl-/1HCO3- exchange
Abstract
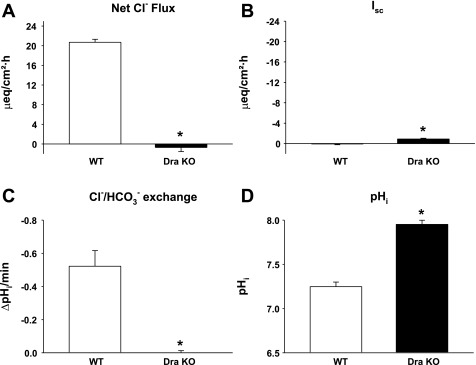
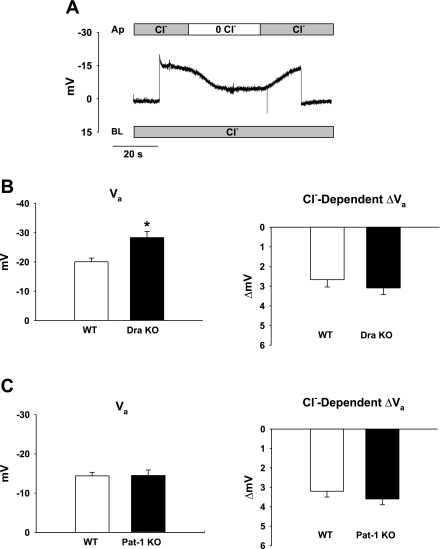
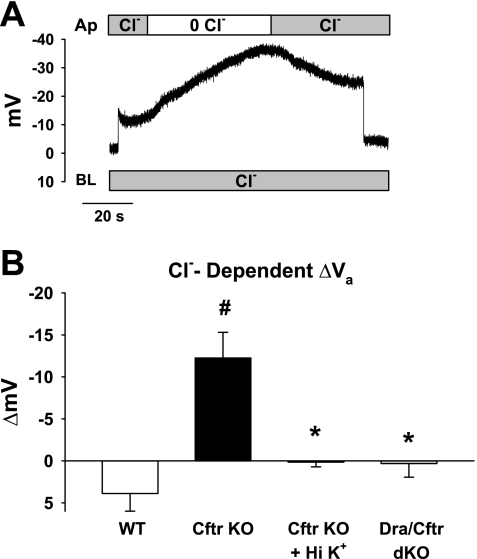
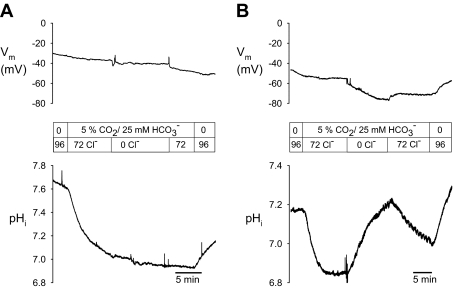
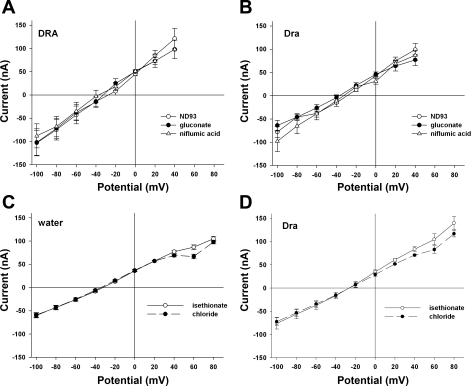
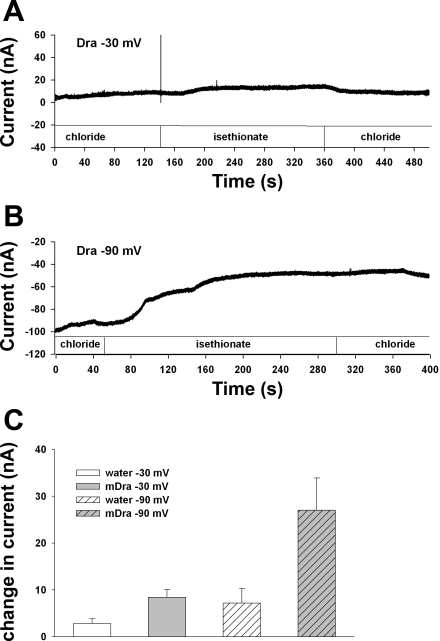
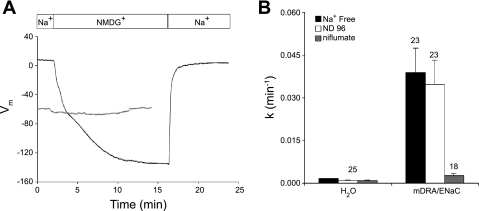
The recent proposal that Dra/Slc26a3 mediates electrogenic 2Cl(-)/1HCO(3)(-) exchange suggests a required revision of classical concepts of electroneutral Cl(-) transport across epithelia such as the intestine. We investigated 1) the effect of endogenous Dra Cl(-)/HCO(3)(-) activity on apical membrane potential (V(a)) of the cecal surface epithelium using wild-type (WT) and knockout (KO) mice; and 2) the electrical properties of Cl(-)/(OH(-))HCO(3)(-) exchange by mouse and human orthologs of Dra expressed in Xenopus oocytes. Ex vivo (36)Cl(-) fluxes and microfluorometry revealed that cecal Cl(-)/HCO(3)(-) exchange was abolished in the Dra KO without concordant changes in short-circuit current. In microelectrode studies, baseline V(a) of Dra KO surface epithelium was slightly hyperpolarized relative to WT but depolarized to the same extent as WT during luminal Cl(-) substitution. Subsequent studies indicated that Cl(-)-dependent V(a) depolarization requires the anion channel Cftr. Oocyte studies demonstrated that Dra-mediated exchange of intracellular Cl(-) for extracellular HCO(3)(-) is accompanied by slow hyperpolarization and a modest outward current, but that the steady-state current-voltage relationship is unaffected by Cl(-) removal or pharmacological blockade. Further, Dra-dependent (36)Cl(-) efflux was voltage-insensitive in oocytes coexpressing the cation channels ENaC or ROMK. We conclude that 1) endogenous Dra and recombinant human/mouse Dra orthologs do not exhibit electrogenic 2Cl(-)/1HCO(3)(-) exchange; and 2) acute induction of Dra Cl(-)/HCO(3)(-) exchange is associated with secondary membrane potential changes representing homeostatic responses. Thus, participation of Dra in coupled NaCl absorption and in uncoupled HCO(3)(-) secretion remains compatible with electroneutrality of these processes, and with the utility of electroneutral transport models for predicting epithelial responses in health and disease.
Figures
References
-
- Alrefai WA, Wen X, Jiang W, Katz JP, Steinbrecher KA, Cohen MB, Williams IR, Dudeja PK, Wu GD. Molecular cloning and promoter analysis of downregulated in adenoma (DRA). Am J Physiol Gastrointest Liver Physiol 293: G923–G934, 2007 - PubMed
-
- Barone S, Amlal H, Xu J, Kujala M, Kere J, Petrovic S, Soleimani M. Differential regulation of basolateral Cl−/HCO3− exchangers SLC26A7 and AE1 in kidney outer medullary collecting duct. J Am Soc Nephrol 15: 2002–2011, 2004 - PubMed
-
- Bukhave K, Rask-Madsen J. Saturation kinetics applied to in vitro effects of low prostaglandin E2 and F2α concentrations on ion transport across human jejunal mucosa. Gastroenterology 78: 32–42, 1980 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials