

A brain-derived MeCP2 complex supports a role for MeCP2 in RNA processing
- PMID: 21070191
- PMCID: PMC3148018
- DOI: 10.1042/BSR20100124
A brain-derived MeCP2 complex supports a role for MeCP2 in RNA processing
Abstract
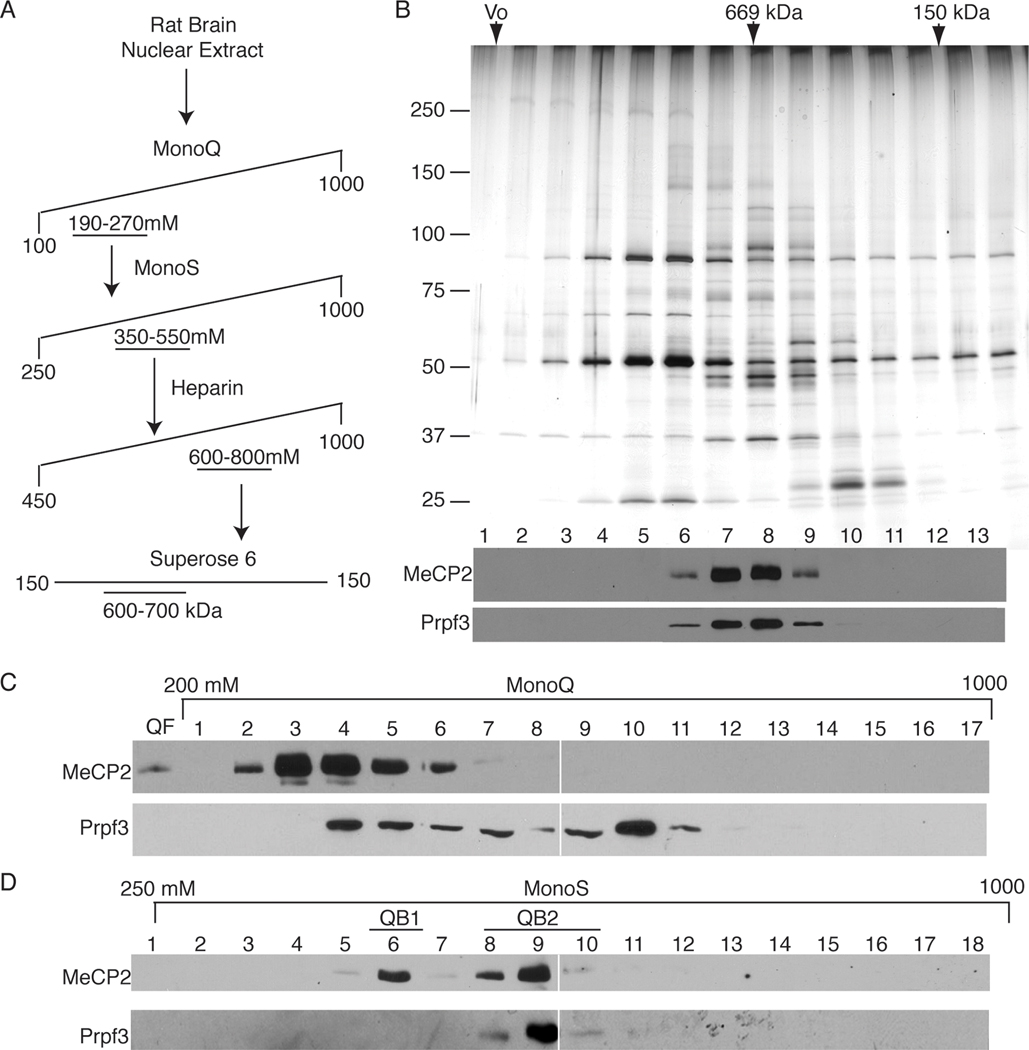
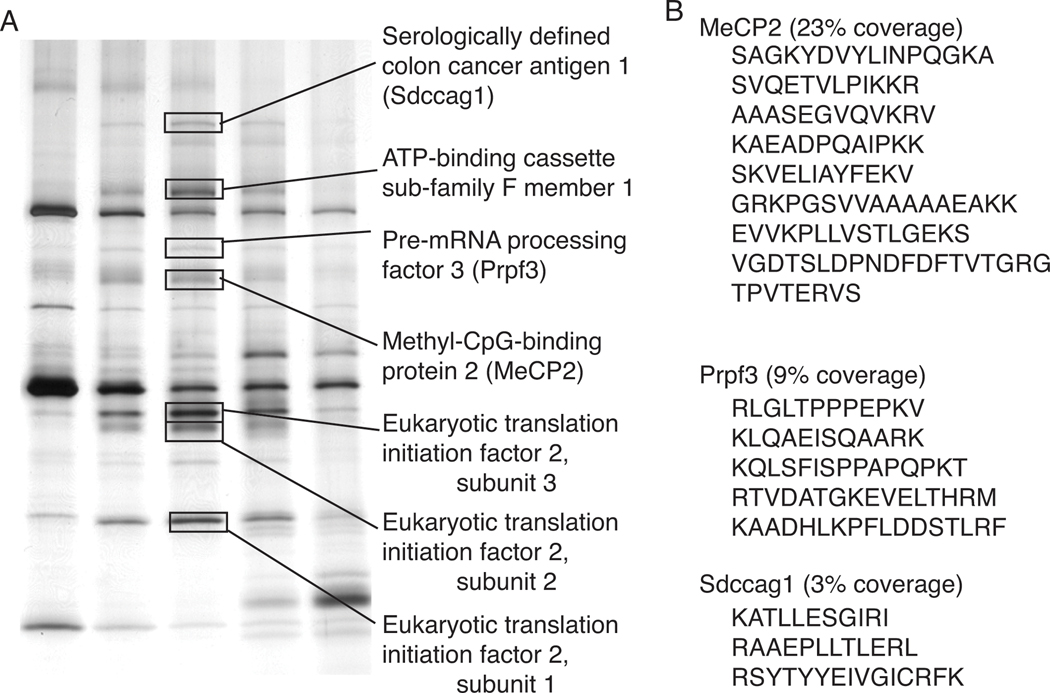
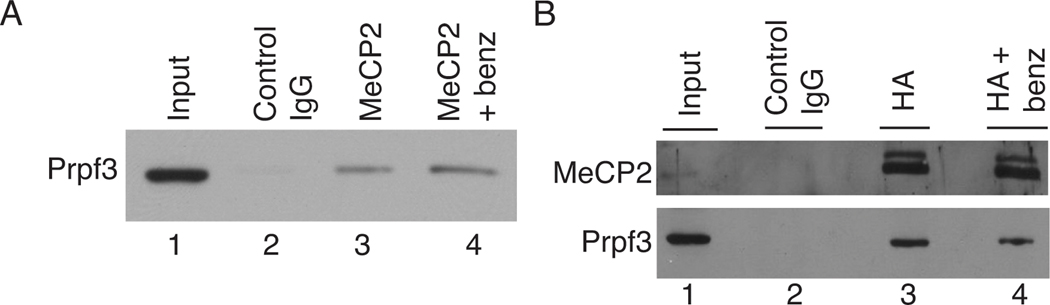
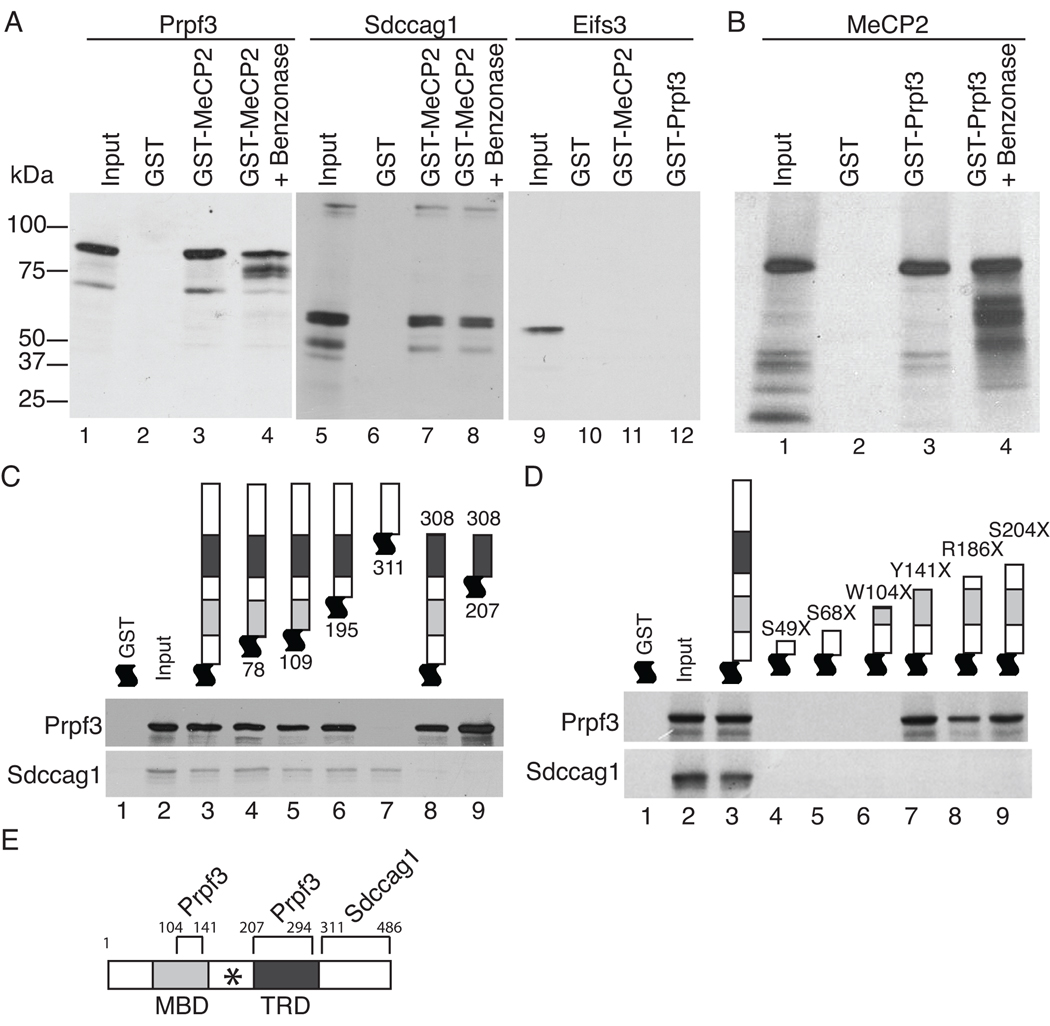
Mutations in MECP2 (methyl-CpG-binding protein 2) are linked to the severe postnatal neurodevelopmental disorder RTT (Rett syndrome). MeCP2 was originally characterized as a transcriptional repressor that preferentially bound methylated DNA; however, recent results indicate MeCP2 is a multifunctional protein. MeCP2 binding is now associated with certain expressed genes and involved in nuclear organization as well, indicating that its gene regulatory function is context-dependent. In addition, MeCP2 is proposed to regulate mRNA splicing and a mouse model for RTT shows aberrant mRNA splicing. To further understand MeCP2 and potential roles in RTT pathogenesis, we have employed a biochemical approach to identify the MeCP2 protein complexes present in the mammalian brain. We show that MeCP2 exists in at least four biochemically distinct pools in the brain and characterize one novel brain-derived MeCP2 complex that contains the splicing factor Prpf3 (pre-mRNA processing factor 3). MeCP2 directly interacts with Prpf3 in vitro and in vivo and many MECP2 RTT truncations disrupt the MeCP2-Prpf3 complex. In addition, MeCP2 and Prpf3 associate in vivo with mRNAs from genes known to be expressed when their promoters are associated with MeCP2. These results support a role for MeCP2 in mRNA biogenesis and suggest an additional mechanism for RTT pathophysiology.
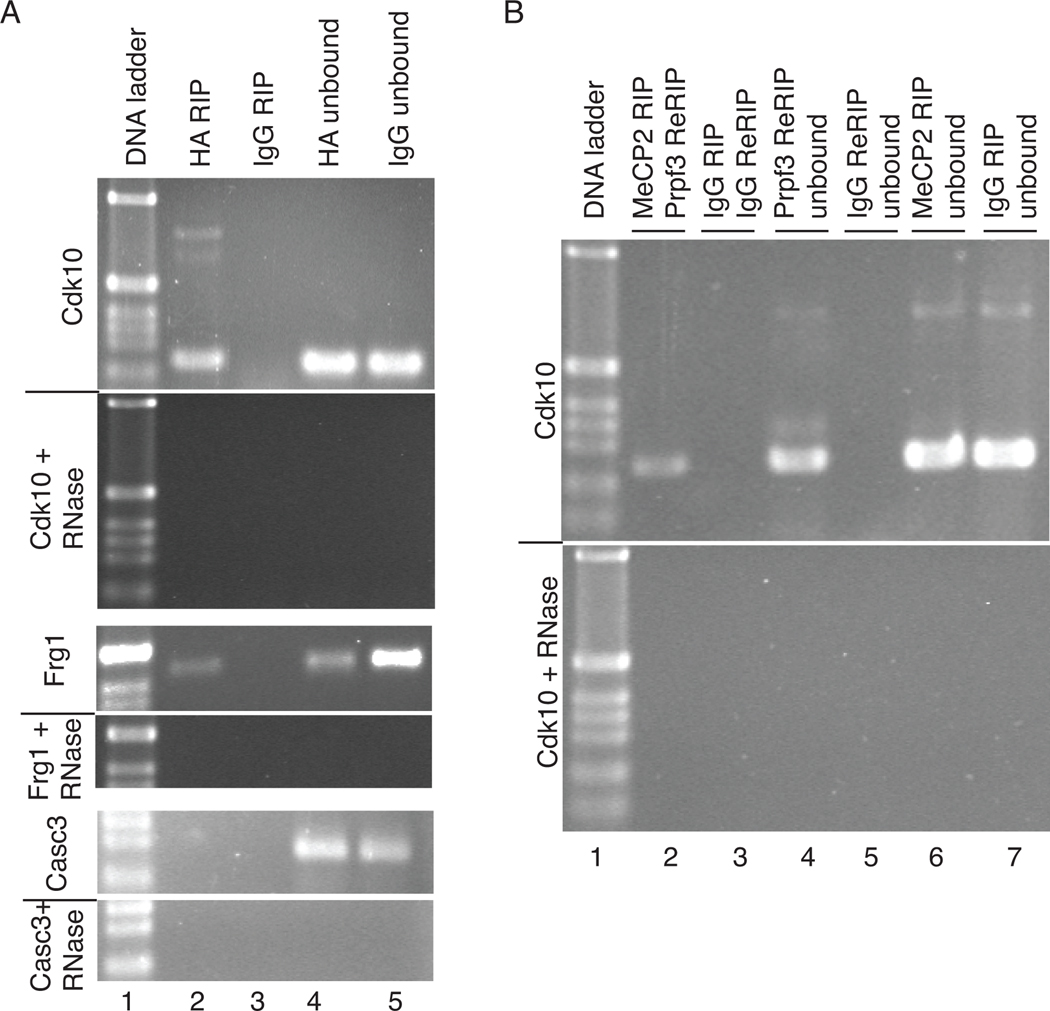
Figures

References
-
- Jones PL, Veenstra GJ, Wade PA, Vermaak D, Kass SU, Landsberger N, Strouboulis J, Wolffe AP. Methylated DNA and MeCP2 recruit histone deacetylase to repress transcription. Nat Genet. 1998;19:187–191. - PubMed
-
- Nan X, Campoy FJ, Bird A. MeCP2 is a transcriptional repressor with abundant binding sites in genomic chromatin. Cell. 1997;88:471–481. - PubMed
-
- Nan X, Ng HH, Johnson CA, Laherty CD, Turner BM, Eisenman RN, Bird A. Transcriptional repression by the methyl-CpG-binding protein MeCP2 involves a histone deacetylase complex. Nature. 1998;393:386–389. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases