

Multiple mRNA decapping enzymes in mammalian cells
- PMID: 21070968
- PMCID: PMC2982215
- DOI: 10.1016/j.molcel.2010.10.010
Multiple mRNA decapping enzymes in mammalian cells
Abstract
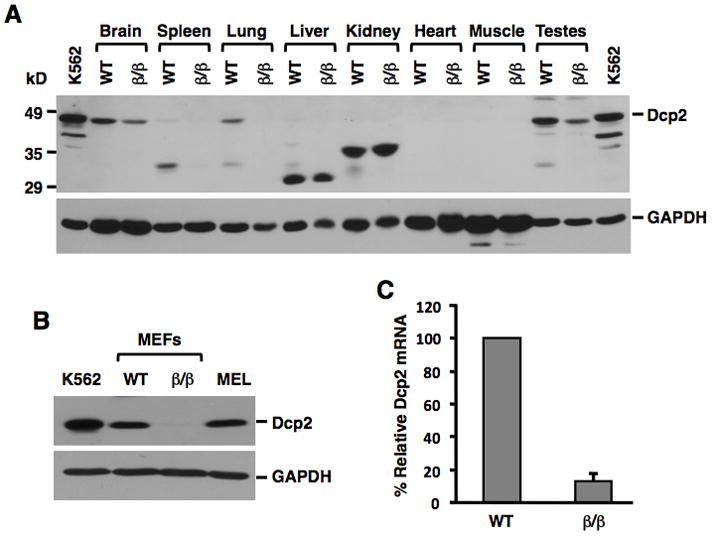
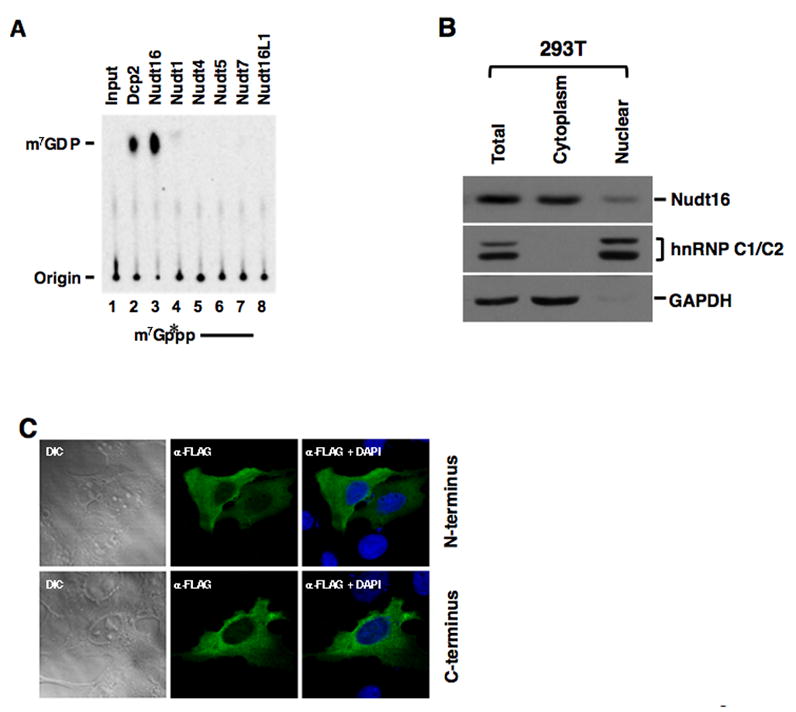
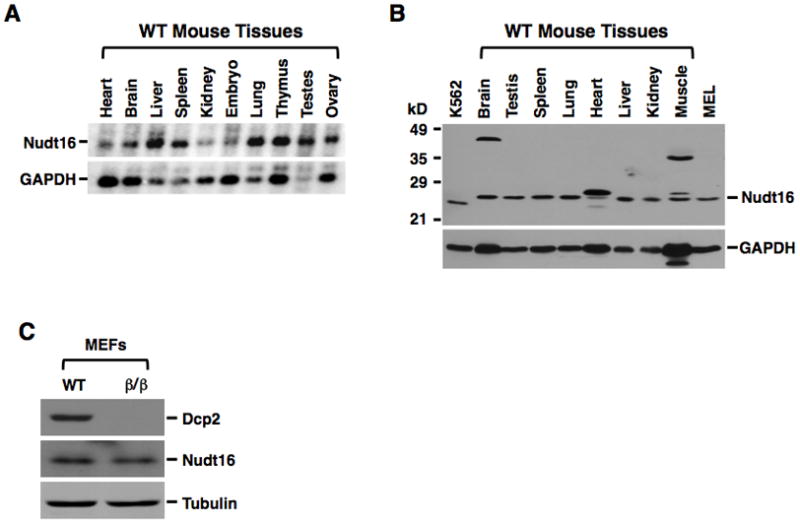
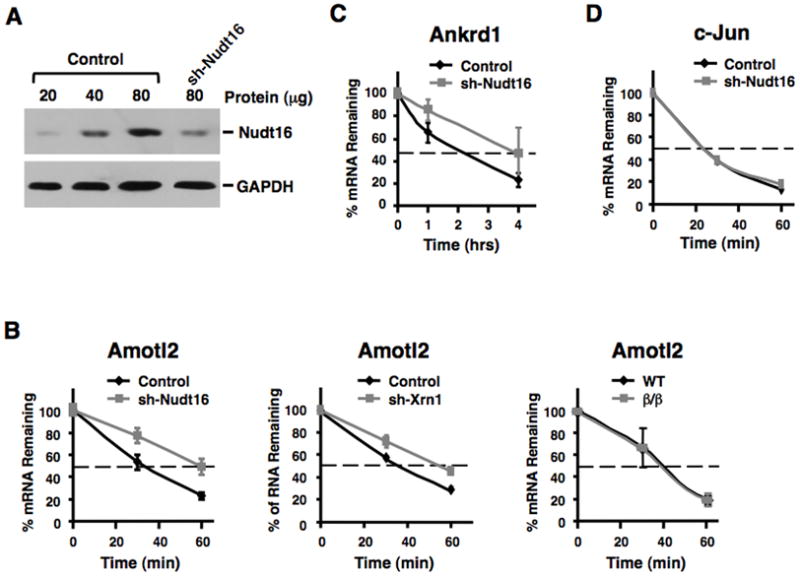
Regulation of RNA degradation plays an important role in the control of gene expression. One mechanism of eukaryotic mRNA decay proceeds through an initial deadenylation followed by 5' end decapping and exonucleolytic decay. Dcp2 is currently believed to be the only cytoplasmic decapping enzyme responsible for decapping of all mRNAs. Here we report that Dcp2 protein modestly contributes to bulk mRNA decay and surprisingly is not detectable in a subset of mouse and human tissues. Consistent with these findings, a hypomorphic knockout of Dcp2 had no adverse consequences in mice. In contrast, the previously reported Xenopus nucleolar decapping enzyme, Nudt16, is an ubiquitous cytoplasmic decapping enzyme in mammalian cells. Like Dcp2, Nudt16 also regulates the stability of a subset of mRNAs including a member of the motin family of proteins involved in angiogenesis, Angiomotin-like 2. These data demonstrate mammalian cells possess multiple mRNA decapping enzymes, including Nudt16 to regulate mRNA turnover.
Copyright © 2010 Elsevier Inc. All rights reserved.
Figures


Comment in
-
Alternate endings: a new story for mRNA decapping.Mol Cell. 2010 Nov 12;40(3):349-50. doi: 10.1016/j.molcel.2010.10.025. Mol Cell. 2010. PMID: 21070961
Similar articles
-
Differential utilization of decapping enzymes in mammalian mRNA decay pathways.RNA. 2011 Mar;17(3):419-28. doi: 10.1261/rna.2439811. Epub 2011 Jan 11. RNA. 2011. PMID: 21224379 Free PMC article.
-
Multiple Nudix family proteins possess mRNA decapping activity.RNA. 2013 Mar;19(3):390-9. doi: 10.1261/rna.037309.112. Epub 2013 Jan 25. RNA. 2013. PMID: 23353937 Free PMC article.
-
Transcript-specific decapping and regulated stability by the human Dcp2 decapping protein.Mol Cell Biol. 2008 Feb;28(3):939-48. doi: 10.1128/MCB.01727-07. Epub 2007 Nov 26. Mol Cell Biol. 2008. PMID: 18039849 Free PMC article.
-
Regulation of mRNA decapping.Wiley Interdiscip Rev RNA. 2010 Sep-Oct;1(2):253-65. doi: 10.1002/wrna.15. Epub 2010 May 6. Wiley Interdiscip Rev RNA. 2010. PMID: 21935889 Review.
-
Structural and functional control of the eukaryotic mRNA decapping machinery.Biochim Biophys Acta. 2013 Jun-Jul;1829(6-7):580-9. doi: 10.1016/j.bbagrm.2012.12.006. Epub 2012 Dec 31. Biochim Biophys Acta. 2013. PMID: 23287066 Free PMC article. Review.
Cited by
-
Eukaryotic RNA 5'-End NAD+ Capping and DeNADding.Trends Cell Biol. 2018 Jun;28(6):454-464. doi: 10.1016/j.tcb.2018.02.005. Epub 2018 Mar 12. Trends Cell Biol. 2018. PMID: 29544676 Free PMC article. Review.
-
Strategies for viral RNA stability: live long and prosper.Trends Genet. 2011 Jul;27(7):286-93. doi: 10.1016/j.tig.2011.04.003. Trends Genet. 2011. PMID: 21640425 Free PMC article. Review.
-
Thermostability, Tunability, and Tenacity of RNA as Rubbery Anionic Polymeric Materials in Nanotechnology and Nanomedicine-Specific Cancer Targeting with Undetectable Toxicity.Chem Rev. 2021 Jul 14;121(13):7398-7467. doi: 10.1021/acs.chemrev.1c00009. Epub 2021 May 26. Chem Rev. 2021. PMID: 34038115 Free PMC article. Review.
-
Dcp2 decapping protein modulates mRNA stability of the critical interferon regulatory factor (IRF) IRF-7.Mol Cell Biol. 2012 Mar;32(6):1164-72. doi: 10.1128/MCB.06328-11. Epub 2012 Jan 17. Mol Cell Biol. 2012. PMID: 22252322 Free PMC article.
-
Mutations in DCPS and EDC3 in autosomal recessive intellectual disability indicate a crucial role for mRNA decapping in neurodevelopment.Hum Mol Genet. 2015 Jun 1;24(11):3172-80. doi: 10.1093/hmg/ddv069. Epub 2015 Feb 20. Hum Mol Genet. 2015. PMID: 25701870 Free PMC article.
References
-
- Badis G, Saveanu C, Fromont-Racine M, Jacquier A. Targeted mRNA degradation by deadenylation-independent decapping. Mol Cell. 2004;15:5–15. - PubMed
-
- Bessman MJ, Frick DN, O'Handley SF. The MutT proteins or “Nudix” hydrolases, a family of versatile, widely distributed, “housecleaning” enzymes. J Biol Chem. 1996;271:25059–25062. - PubMed
-
- Bratt A, Birot O, Sinha I, Veitonmaki N, Aase K, Ernkvist M, Holmgren L. Angiomotin regulates endothelial cell-cell junctions and cell motility. J Biol Chem. 2005;280:34859–34869. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
