

A stress-responsive system for mitochondrial protein degradation
- PMID: 21070972
- PMCID: PMC2998070
- DOI: 10.1016/j.molcel.2010.10.021
A stress-responsive system for mitochondrial protein degradation
Abstract
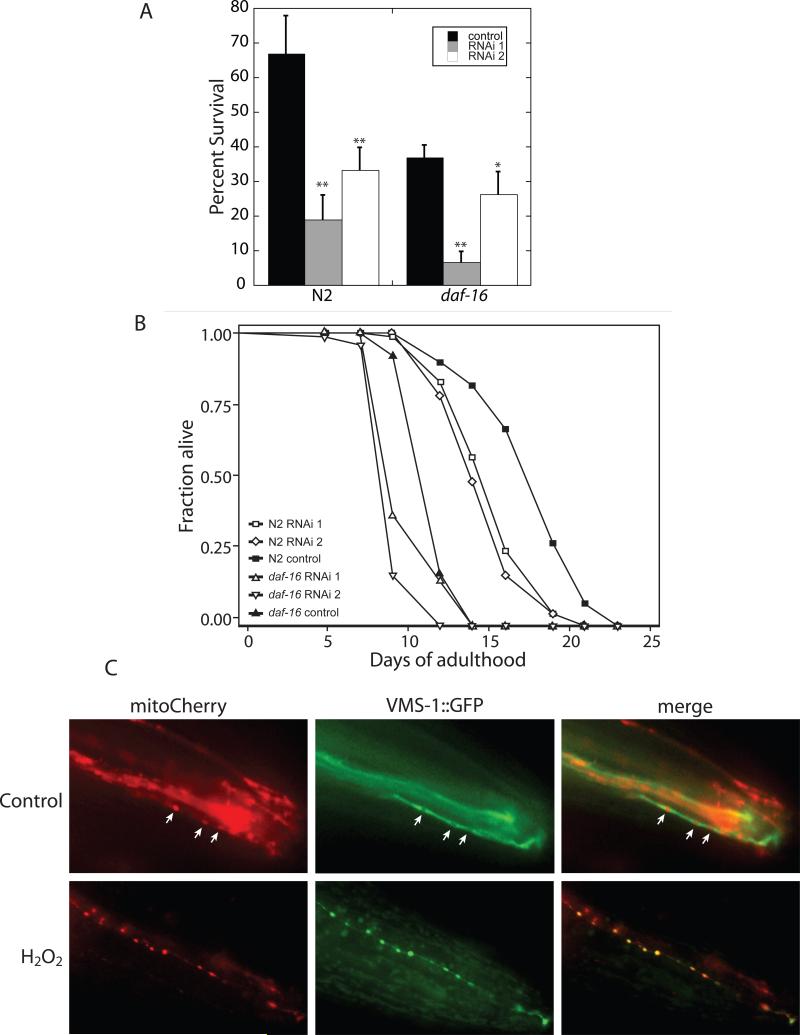
We show that Ydr049 (renamed VCP/Cdc48-associated mitochondrial stress-responsive--Vms1), a member of an unstudied pan-eukaryotic protein family, translocates from the cytosol to mitochondria upon mitochondrial stress. Cells lacking Vms1 show progressive mitochondrial failure, hypersensitivity to oxidative stress, and decreased chronological life span. Both yeast and mammalian Vms1 stably interact with Cdc48/VCP/p97, a component of the ubiquitin/proteasome system with a well-defined role in endoplasmic reticulum-associated protein degradation (ERAD), wherein misfolded ER proteins are degraded in the cytosol. We show that oxidative stress triggers mitochondrial localization of Cdc48 and this is dependent on Vms1. When this system is impaired by mutation of Vms1, ubiquitin-dependent mitochondrial protein degradation, mitochondrial respiratory function, and cell viability are compromised. We demonstrate that Vms1 is a required component of an evolutionarily conserved system for mitochondrial protein degradation, which is necessary to maintain mitochondrial, cellular, and organismal viability.
Copyright © 2010 Elsevier Inc. All rights reserved.
Figures






Comment in
-
Stressed-out mitochondria get MAD.Cell Metab. 2010 Dec 1;12(6):559-60. doi: 10.1016/j.cmet.2010.11.018. Cell Metab. 2010. PMID: 21109188 Free PMC article.
References
-
- Bays NW, Hampton RY. Cdc48-Ufd1-Npl4: stuck in the middle with Ub. Curr Biol. 2002;12:R366–371. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
