

Efficient preparation of internally modified single-molecule constructs using nicking enzymes
- PMID: 21071409
- PMCID: PMC3035433
- DOI: 10.1093/nar/gkq1004
Efficient preparation of internally modified single-molecule constructs using nicking enzymes
Abstract
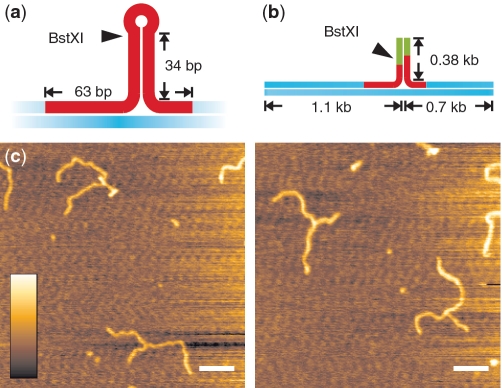
Investigations of enzymes involved in DNA metabolism have strongly benefited from the establishment of single molecule techniques. These experiments frequently require elaborate DNA substrates, which carry chemical labels or nucleic acid tertiary structures. Preparing such constructs often represents a technical challenge: long modified DNA molecules are usually produced via multi-step processes, involving low efficiency intermolecular ligations of several fragments. Here, we show how long stretches of DNA (>50 bp) can be modified using nicking enzymes to produce complex DNA constructs. Multiple different chemical and structural modifications can be placed internally along DNA, in a specific and precise manner. Furthermore, the nicks created can be resealed efficiently yielding intact molecules, whose mechanical properties are preserved. Additionally, the same strategy is applied to obtain long single-strand overhangs subsequently used for efficient ligation of ss- to dsDNA molecules. This technique offers promise for a wide range of applications, in particular single-molecule experiments, where frequently multiple internal DNA modifications are required.
Figures





Similar articles
-
T4 endonuclease VII resolves cruciform DNA with nick and counter-nick and its activity is directed by local nucleotide sequence.J Mol Biol. 1992 Feb 5;223(3):607-15. doi: 10.1016/0022-2836(92)90977-r. J Mol Biol. 1992. PMID: 1542108
-
[Investigation of site-specific DNA binding with nicking endonuclease Nt.BspD6I at single molecule level by atomic force microscopy].Biofizika. 2012 May-Jun;57(3):432-6. Biofizika. 2012. PMID: 22873066 Russian.
-
Initiation of translocation by Type I restriction-modification enzymes is associated with a short DNA extrusion.Nucleic Acids Res. 2004 Dec 14;32(22):6540-7. doi: 10.1093/nar/gkh999. Print 2004. Nucleic Acids Res. 2004. PMID: 15598825 Free PMC article.
-
Novel drug discovery and molecular biological methods, via DNA, RNA and protein changes using structure-function transitions: Transitional structural chemogenomics, transitional structural chemoproteomics and novel multi-stranded nucleic acid microarray.Med Hypotheses. 2006;67(5):1099-114. doi: 10.1016/j.mehy.2006.05.020. Epub 2006 Jul 7. Med Hypotheses. 2006. PMID: 16828979 Review.
-
Single-molecule manipulation quantification of site-specific DNA binding.Curr Opin Chem Biol. 2019 Dec;53:106-117. doi: 10.1016/j.cbpa.2019.08.006. Epub 2019 Oct 31. Curr Opin Chem Biol. 2019. PMID: 31677535 Review.
Cited by
-
Locked nucleic acid oligomers as handles for single molecule manipulation.Nucleic Acids Res. 2014 Oct 29;42(19):e150. doi: 10.1093/nar/gku760. Epub 2014 Aug 26. Nucleic Acids Res. 2014. PMID: 25159617 Free PMC article.
-
Human HELB is a processive motor protein that catalyzes RPA clearance from single-stranded DNA.Proc Natl Acad Sci U S A. 2022 Apr 12;119(15):e2112376119. doi: 10.1073/pnas.2112376119. Epub 2022 Apr 6. Proc Natl Acad Sci U S A. 2022. PMID: 35385349 Free PMC article.
-
Unwinding forward and sliding back: an intermittent unwinding mode of the BLM helicase.Nucleic Acids Res. 2015 Apr 20;43(7):3736-46. doi: 10.1093/nar/gkv209. Epub 2015 Mar 12. Nucleic Acids Res. 2015. PMID: 25765643 Free PMC article.
-
Nuclease activity of Saccharomyces cerevisiae Dna2 inhibits its potent DNA helicase activity.Proc Natl Acad Sci U S A. 2013 May 28;110(22):E1992-2001. doi: 10.1073/pnas.1300390110. Epub 2013 May 13. Proc Natl Acad Sci U S A. 2013. PMID: 23671118 Free PMC article.
-
Multiple RPAs make WRN syndrome protein a superhelicase.Nucleic Acids Res. 2018 May 18;46(9):4689-4698. doi: 10.1093/nar/gky272. Nucleic Acids Res. 2018. PMID: 29668972 Free PMC article.
References
-
- Bustamante C, Marko JF, Siggia ED, Smith S. Entropic elasticity of lambda-phage DNA. Science. 1994;265:1599–1600. - PubMed
-
- Seidel R, Dekker C. Single-molecule studies of nucleic acid motors. Curr. Opin. Struct. Biol. 2007;17:80–86. - PubMed
-
- Kapanidis AN, Strick T. Biology, one molecule at a time. Trends Biochem. Sci. 2009;34:234–243. - PubMed
