

Resveratrol blocks Akt activation in angiotensin II- or EGF-stimulated vascular smooth muscle cells in a redox-independent manner
- PMID: 21071431
- PMCID: PMC3058730
- DOI: 10.1093/cvr/cvq355
Resveratrol blocks Akt activation in angiotensin II- or EGF-stimulated vascular smooth muscle cells in a redox-independent manner
Abstract
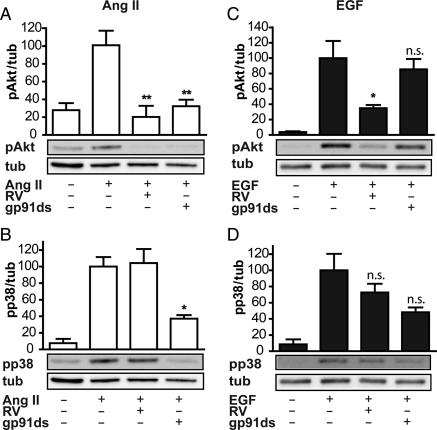
Aims: Resveratrol (RV), an antioxidant, inhibits angiotensin II (Ang II)-induced hypertrophy and Ang II- or epidermal growth factor (EGF)-induced Akt phosphorylation in rat vascular smooth muscle cells (VSMCs). Both signalling pathways are reported to utilize reactive oxygen species (ROS). The aim of this study was to show whether RV reduces the ROS level in Ang II- or EGF-activated VSMCs and whether reduction of ROS causes the impeded signalling towards Akt in the presence of RV.
Methods and results: We show here that RV reduces intracellular ROS and extracellular H₂O₂ release from VSMCs as measured using 2',7'-dichlorodihydrofluorescein-diacetate and Amplex Red™. Since NADPH oxidases (Nox) 1 and 4 are major ROS sources in VSMCs, we examined their need for Akt phosphorylation in response to Ang II or EGF. Experiments using the blocking peptide gp91ds-tat verified a role for Nox1 in Ang II signalling towards Akt, but excluded a role for Nox1 in the respective EGF signalling. A small interfering RNA-mediated knock-down of Nox4 showed that Nox4 was not required for Ang II- or EGF-induced Akt phosphorylation. Use of the flavoprotein inhibitor diphenyleneiodonium, N-acetyl-cysteine, and non-antioxidant RV derivatives revealed that the antioxidant capacity of RV is not required for the inhibition of Akt phosphorylation, in both rat and human VSMCs.
Conclusion: Thus, although RV acts as an antioxidant, the antihypertrophic response of RV in VSMCs and the signalling downstream of the EGF receptor towards Akt seem to be largely redox independent.
Figures





References
-
- Schmitt CA, Dirsch VM. Modulation of endothelial nitric oxide by plant-derived products. Nitric Oxide. 2009;21:77–91. doi:10.1016/j.niox.2009.05.006. - DOI - PubMed
-
- Opie LH, Lecour S. The red wine hypothesis: from concepts to protective signalling molecules. Eur Heart J. 2007;28:1683–1693. doi:10.1093/eurheartj/ehm149. - DOI - PubMed
-
- Leifert WR, Abeywardena MY. Cardioprotective actions of grape polyphenols. Nutr Res. 2008;28:729–737. doi:10.1016/j.nutres.2008.08.007. - DOI - PubMed
-
- Haider UG, Sorescu D, Griendling KK, Vollmar AM, Dirsch VM. Resveratrol suppresses angiotensin II-induced Akt/protein kinase B and p70 S6 kinase phosphorylation and subsequent hypertrophy in rat aortic smooth muscle cells. Mol Pharmacol. 2002;62:772–777. doi:10.1124/mol.62.4.772. - DOI - PubMed
-
- Haider UG, Roos TU, Kontaridis MI, Neel BG, Sorescu D, Griendling KK, et al. Resveratrol inhibits angiotensin II- and epidermal growth factor-mediated Akt activation: role of Gab1 and Shp2. Mol Pharmacol. 2005;68:41–48. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
