

Glucose availability in hypoxia regulates the selection of chronic myeloid leukemia progenitor subsets with different resistance to imatinib-mesylate
- PMID: 21071498
- PMCID: PMC3031687
- DOI: 10.3324/haematol.2010.029082
Glucose availability in hypoxia regulates the selection of chronic myeloid leukemia progenitor subsets with different resistance to imatinib-mesylate
Abstract
Background: Incubation of chronic myeloid leukemia cells in hypoxia inhibits growth and selects BCR/Abl-independent cells with stem cell properties which are refractory to imatinib-mesylate. This study aimed to characterize the relationship of this refractoriness with glucose availability in the environment.
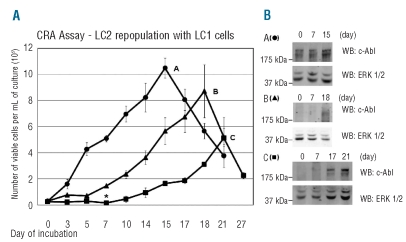
Design and methods: K562 or primary chronic myeloid leukemia cells were cultured at 0.1% O(2), different cell densities and glucose concentrations. The stem and progenitor cell potential of these cultures at different times of incubation in relation to BCR/Abl(protein) expression and sensitivity to imatinib-mesylate was explored by transferring cells to growth-permissive secondary cultures in normoxia, according to the Culture-Repopulating Ability assay methodology.
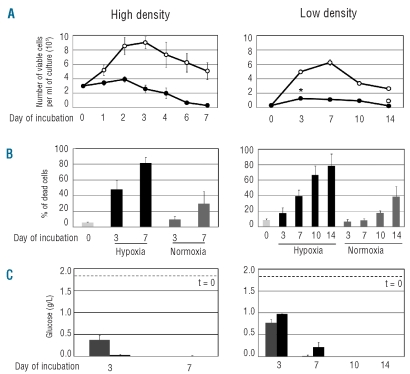
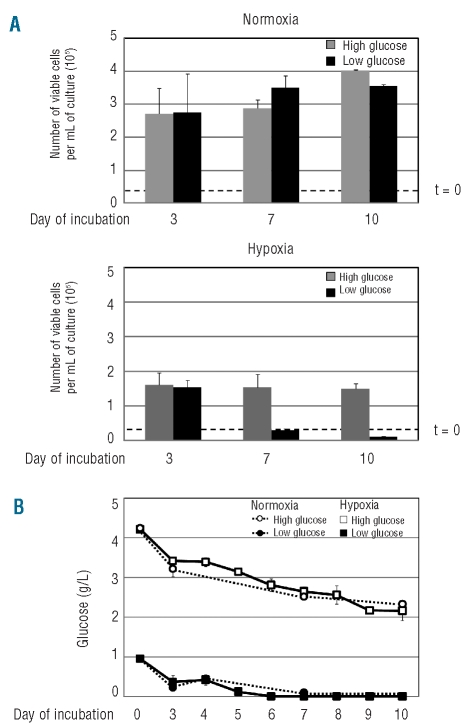
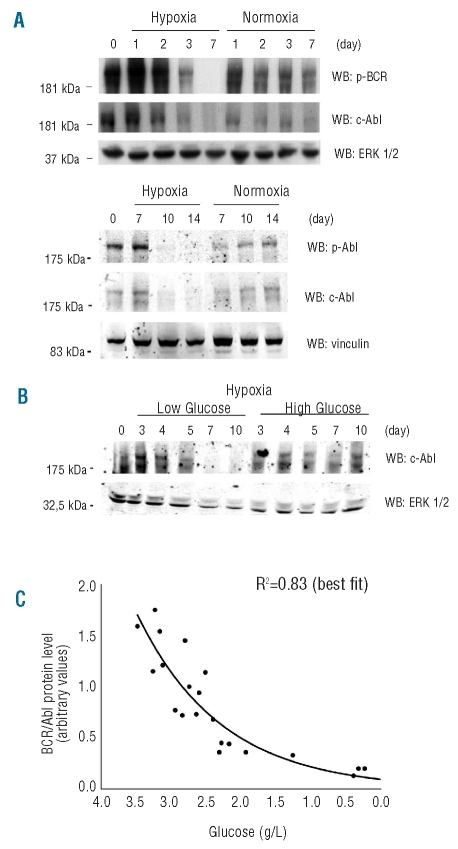
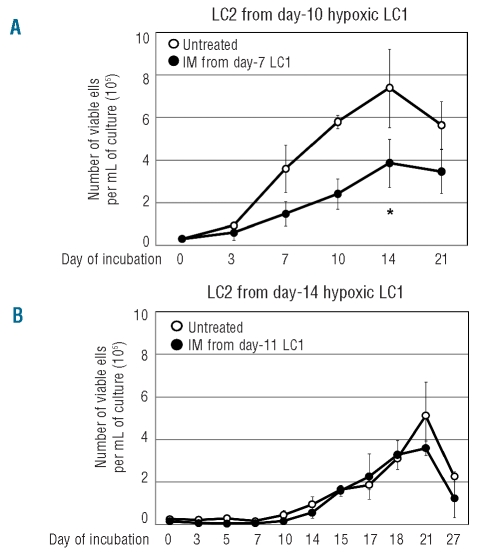
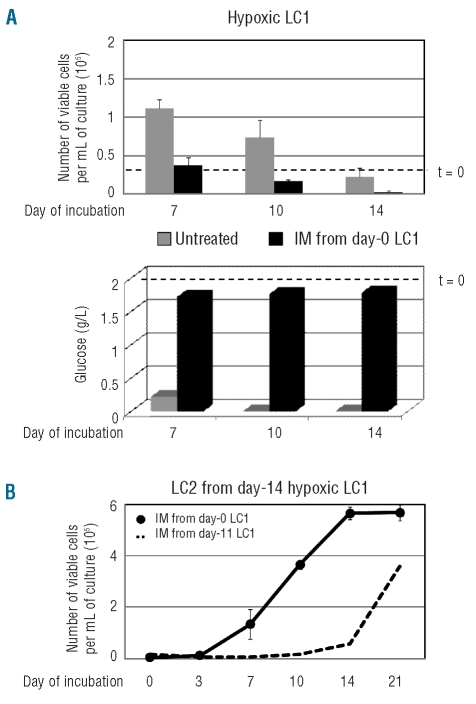
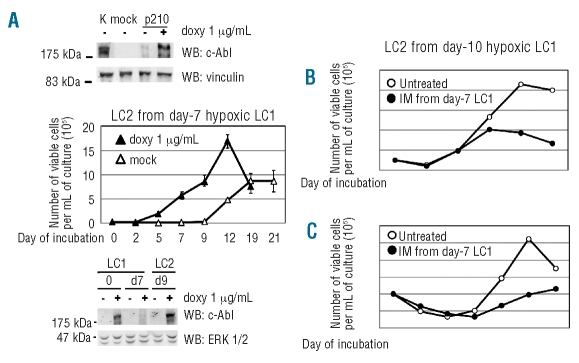
Results: Hypoxia-resistant cells maintained BCR/Abl(protein) expression until glucose was no longer available in primary hypoxic cultures, where glucose availability appeared to regulate cell number and the balance between the enrichment of cells with kinetic properties typical of stem or progenitor cells. Cells surviving merely hypoxic conditions were, upon transfer to secondary cultures, immediately available for numerical expansion due to the maintained BCR/Abl(protein) expression, and were consequently sensitive to imatinib-mesylate. Instead, BCR/Abl(protein)-negative cells selected in primary cultures under oxygen/glucose shortage underwent a delayed numerical expansion in secondary cultures, which was completely refractory to imatinib-mesylate. Cells with the latter properties were also found in primary chronic myeloid leukemia explants.
Conclusions: Glucose shortage in hypoxia was shown to represent the condition selecting BCR/Abl(protein)-negative cells refractory to imatinib-mesylate from either chronic myeloid leukemia lines or patients. These cells, exhibiting stem cell properties in vitro, are metabolically suited to home to stem cell niches in vivo and so may represent the chronic myeloid leukemia cell subset responsible for minimal residual disease.
Figures
References
-
- Giuntoli S, Rovida E, Gozzini A, Barbetti V, Cipolleschi MG, Olivotto M, Dello Sbarba P. Severe hypoxia defines heterogeneity and selects highly immature progenitors within clonal erythroleukemia cells. Stem Cells. 2007;25(5):1119–25. - PubMed
-
- Giuntoli S, Rovida E, Barbetti V, Cipolleschi MG, Olivotto M, Dello Sbarba P. Hypoxia suppresses BCR/Abl and selects Imatinib-insensitive progenitors within clonal CML population. Leukemia. 2006;20(7):1291–3. - PubMed
-
- Dello Sbarba P, Cipolleschi MG, Olivotto M. Hemopoietic progenitor cells are sensitive to the cytostatic effect of pyruvate. Exp Hematol. 1987;15(2):137–42. - PubMed
-
- Cipolleschi MG, Dello Sbarba P, Olivotto M. The role of hypoxia in the maintenance of hematopoietic stem cells. Blood. 1993;82(7):2031–7. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
