

Calcitonin has a vasopressin-like effect on aquaporin-2 trafficking and urinary concentration
- PMID: 21071524
- PMCID: PMC3014035
- DOI: 10.1681/ASN.2009121267
Calcitonin has a vasopressin-like effect on aquaporin-2 trafficking and urinary concentration
Abstract
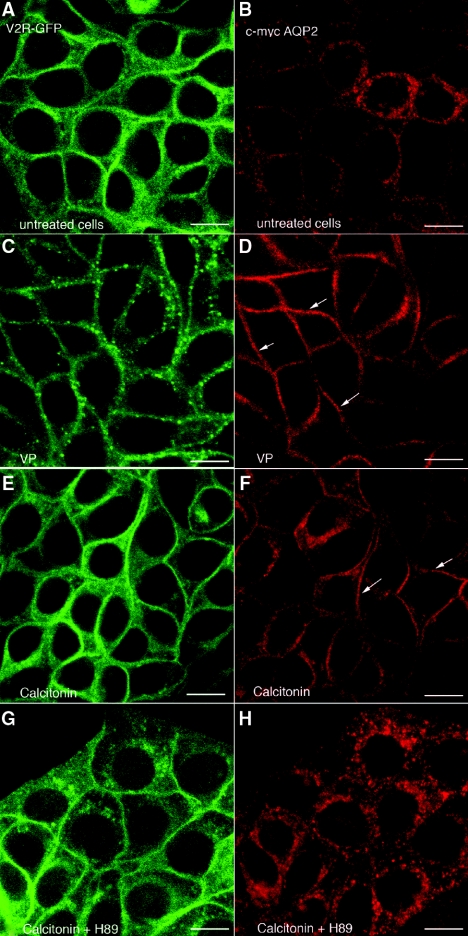
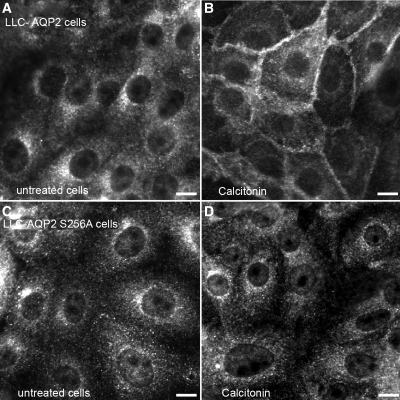
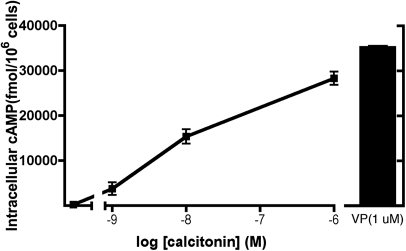
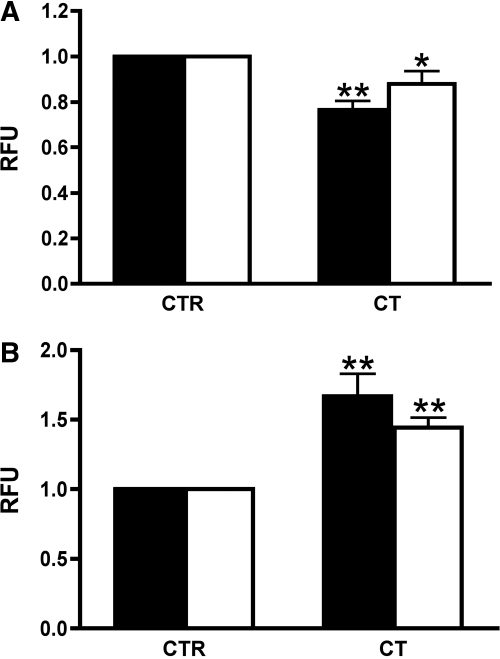
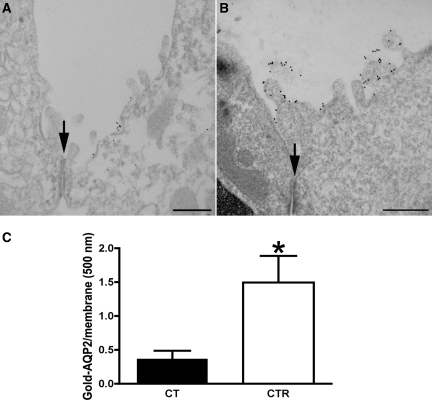
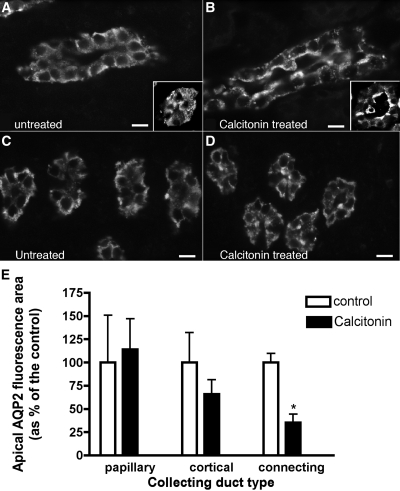
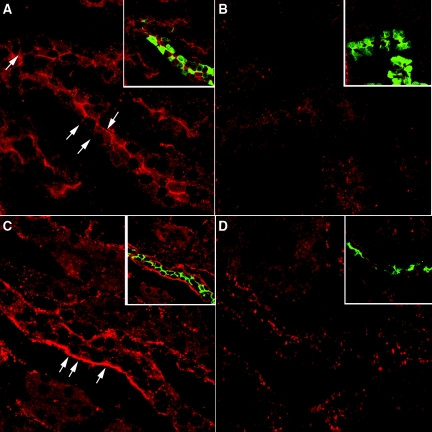
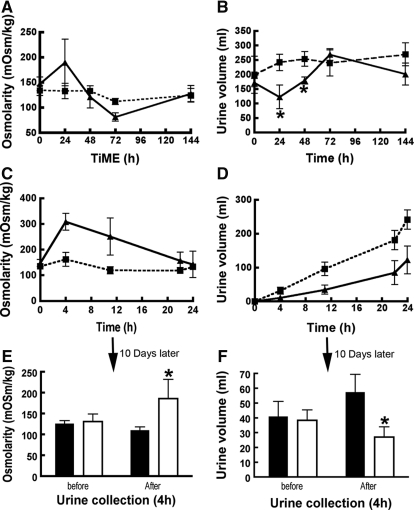
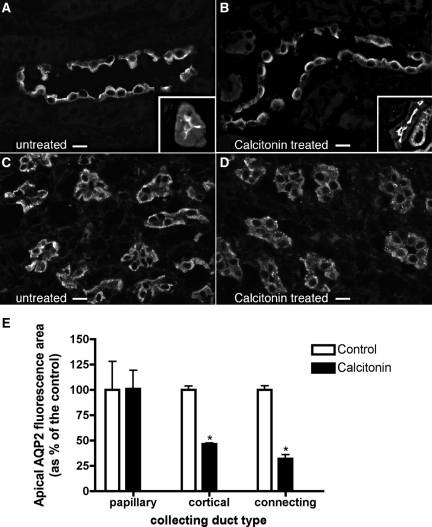
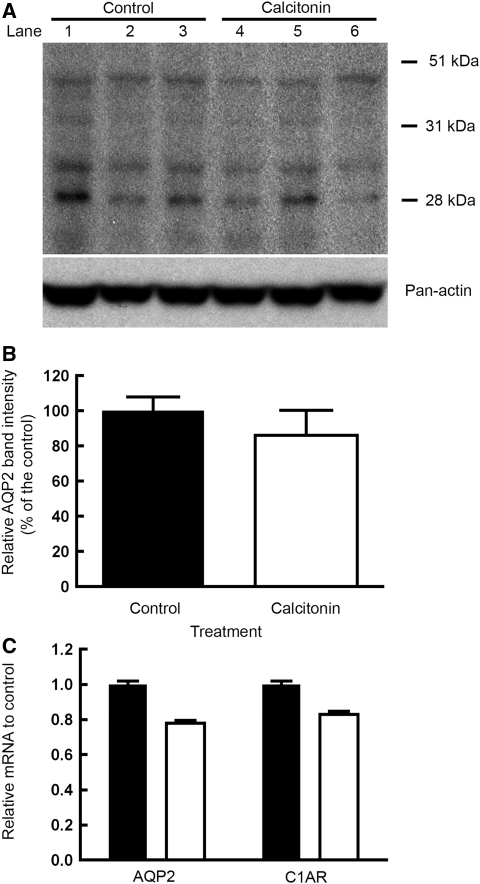
The most common cause of hereditary nephrogenic diabetes insipidus is a nonfunctional vasopressin (VP) receptor type 2 (V2R). Calcitonin, another ligand of G-protein-coupled receptors, has a VP-like effect on electrolytes and water reabsorption, suggesting that it may affect AQP2 trafficking. Here, calcitonin increased intracellular cAMP and stimulated the membrane accumulation of AQP2 in LLC-PK1 cells. Pharmacologic inhibition of protein kinase A (PKA) and deficiency of a critical PKA phosphorylation site on AQP2 both prevented calcitonin-induced membrane accumulation of AQP2. Fluorescence assays showed that calcitonin led to a 70% increase in exocytosis and a 20% decrease in endocytosis of AQP2. Immunostaining of rat kidney slices demonstrated that calcitonin induced a significant redistribution of AQP2 to the apical membrane of principal cells in cortical collecting ducts and connecting segments but not in the inner stripe or inner medulla. Calcitonin-treated VP-deficient Brattleboro rats had a reduced urine flow and two-fold higher urine osmolality during the first 12 hours of treatment compared with control groups. Although this VP-like effect of calcitonin diminished over the following 72 hours, the tachyphylaxis was reversible. Taken together, these data show that calcitonin induces cAMP-dependent AQP2 trafficking in cortical collecting and connecting tubules in parallel with an increase in urine concentration. This suggests that calcitonin has a potential therapeutic use in nephrogenic diabetes insipidus.
Figures
References
-
- Bichet DG, Razi M, Arthus MF, Lonergan M, Tittley P, Smiley RK, Rock G, Hirsch DJ: Epinephrine and dDAVP administration in patients with congenital nephrogenic diabetes insipidus: Evidence for a pre-cyclic AMP V2 receptor defective mechanism. Kidney Int 36: 859–866, 1989 - PubMed
-
- Canfield MC, Tamarappoo BK, Moses AM, Verkman AS, Holtzman EJ: Identification and characterization of aquaporin-2 water channel mutations causing nephrogenic diabetes insipidus with partial vasopressin response. Hum Mol Genet 6: 1865–1871, 1997 - PubMed
-
- Vargas-Poussou R, Forestier L, Dautzenberg MD, Niaudet P, Dechaux M, Antignac C: Mutations in the vasopressin V2 receptor and aquaporin-2 genes in 12 families with congenital nephrogenic diabetes insipidus. J Am Soc Nephrol 8: 1855–1862, 1997 - PubMed
-
- Brown D: The ins and outs of aquaporin-2 trafficking. Am J Physiol Renal Physiol 284: F893–F901, 2003 - PubMed
-
- Deen PM, Verdijk MA, Knoers NV, Wieringa B, Monnens LA, van Os CH, van Oost BA: Requirement of human renal water channel aquaporin-2 for vasopressin-dependent concentration of urine. Science 264: 92–95, 1994 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
