

Manipulating broad-spectrum disease resistance by suppressing pathogen-induced auxin accumulation in rice
- PMID: 21071600
- PMCID: PMC3075746
- DOI: 10.1104/pp.110.163774
Manipulating broad-spectrum disease resistance by suppressing pathogen-induced auxin accumulation in rice
Abstract
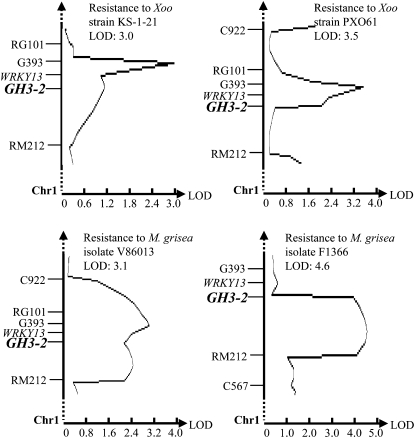
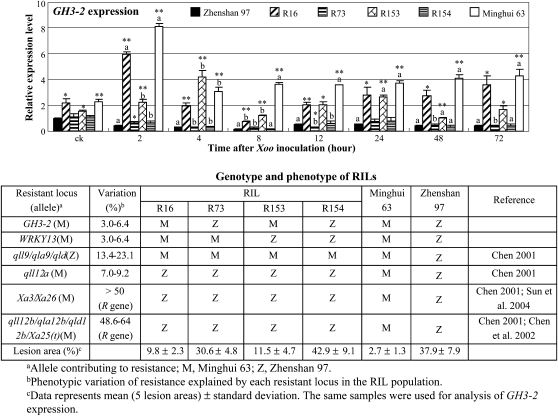
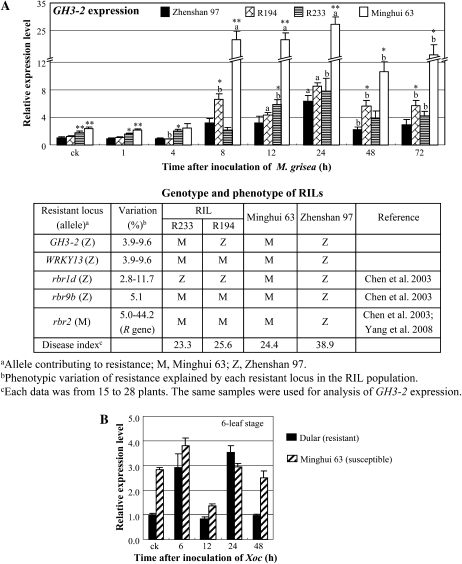
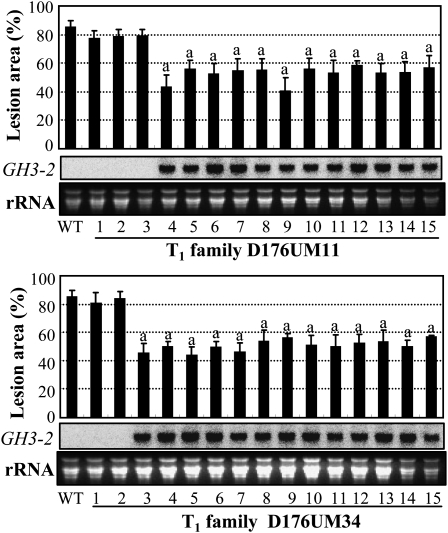
Breeding crops with the quality of broad-spectrum disease resistance using genetic resources is one of the principal goals of crop improvement. However, the molecular mechanism of broad-spectrum resistance remains largely unknown. Here, we show that GH3-2, encoding an indole-3-acetic acid (IAA)-amido synthetase, mediates a broad-spectrum resistance to bacterial Xanthomonas oryzae pv oryzae and Xanthomonas oryzae pv oryzicola and fungal Magnaporthe grisea in rice (Oryza sativa). IAA, the major form of auxin in rice, results in rice more vulnerable to the invasion of different types of pathogens, which is at least partly due to IAA-induced loosening of the cell wall, the natural protective barrier of plant cells to invaders. X. oryzae pv oryzae, X. oryzae pv oryzicola, and M. grisea secrete IAA, which, in turn, may induce rice to synthesize its own IAA at the infection site. IAA induces the production of expansins, the cell wall-loosening proteins, and makes rice vulnerable to pathogens. GH3-2 is likely contributing to a minor quantitative trait locus for broad-spectrum resistance. Activation of GH3-2 inactivates IAA by catalyzing the formation of an IAA-amino acid conjugate, which results in the suppression of expansin genes. Thus, GH3-2 mediates basal resistance by suppressing pathogen-induced IAA accumulation. It is expected that, regulated by a pathogen-induced strong promoter, GH3-2 alone may be used for breeding rice with a broad-spectrum disease resistance.
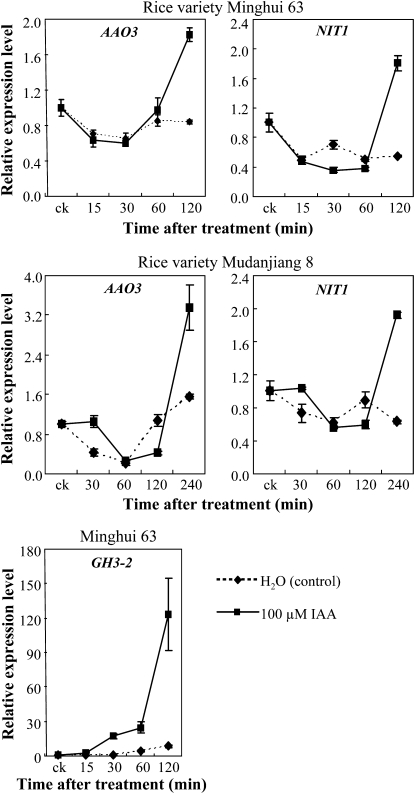
Figures
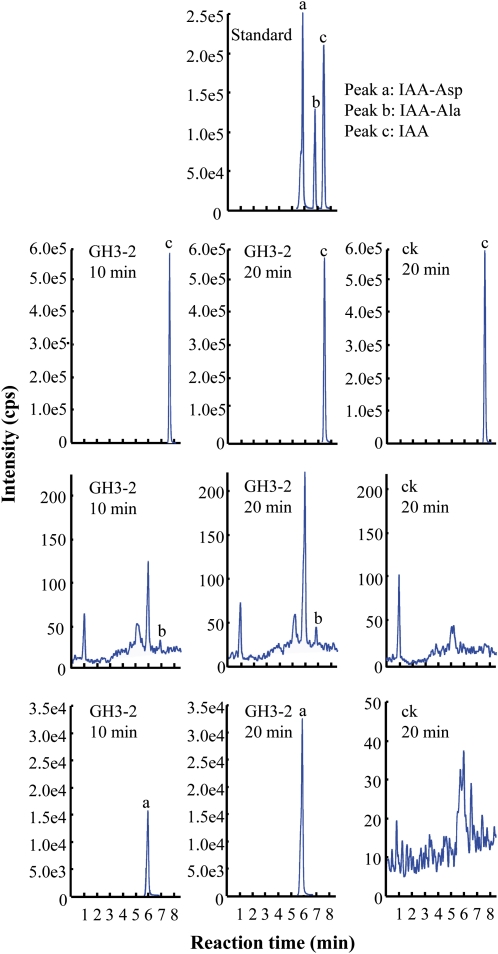
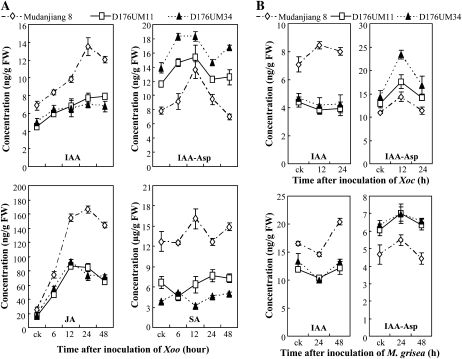
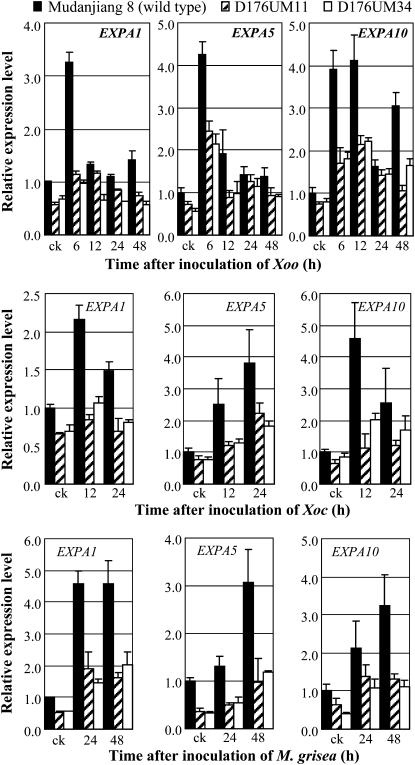
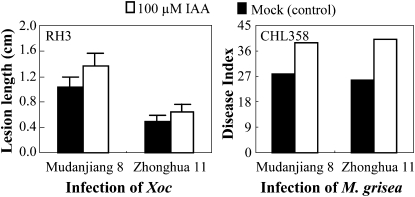

References
-
- Ballini E, Morel JB, Droc G, Price A, Courtois B, Notteghem JL, Tharreau D. (2008) A genome-wide meta-analysis of rice blast resistance genes and quantitative trait loci provides new insights into partial and complete resistance. Mol Plant Microbe Interact 21: 859–868 - PubMed
-
- Bari R, Jones JD. (2009) Role of plant hormones in plant defence responses. Plant Mol Biol 69: 473–488 - PubMed
-
- Cernadas RA, Benedetti CE. (2009) Role of auxin and gibberellin in citrus canker development and in the transcriptional control of cell-wall remodeling genes modulated by Xanthomonas axonopodis pv. citri. Plant Sci 177: 190–195
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
