

Simple ecological trade-offs give rise to emergent cross-ecosystem distributions of a coral reef fish
- PMID: 21072542
- PMCID: PMC3015207
- DOI: 10.1007/s00442-010-1833-8
Simple ecological trade-offs give rise to emergent cross-ecosystem distributions of a coral reef fish
Abstract
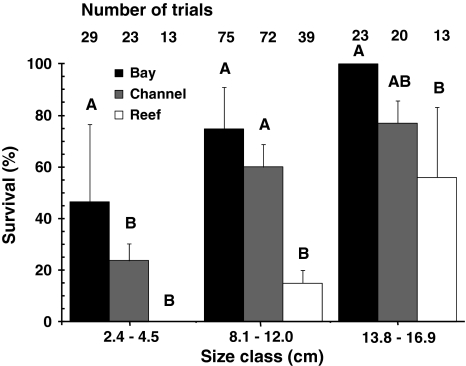
Ecosystems are intricately linked by the flow of organisms across their boundaries, and such connectivity can be essential to the structure and function of the linked ecosystems. For example, many coral reef fish populations are maintained by the movement of individuals from spatially segregated juvenile habitats (i.e., nurseries, such as mangroves and seagrass beds) to areas preferred by adults. It is presumed that nursery habitats provide for faster growth (higher food availability) and/or low predation risk for juveniles, but empirical data supporting this hypothesis is surprisingly lacking for coral reef fishes. Here, we investigate potential mechanisms (growth, predation risk, and reproductive investment) that give rise to the distribution patterns of a common Caribbean reef fish species, Haemulon flavolineatum (French grunt). Adults were primarily found on coral reefs, whereas juvenile fish only occurred in non-reef habitats. Contrary to our initial expectations, analysis of length-at-age revealed that growth rates were highest on coral reefs and not within nursery habitats. Survival rates in tethering trials were 0% for small juvenile fish transplanted to coral reefs and 24-47% in the nurseries. As fish grew, survival rates on coral reefs approached those in non-reef habitats (56 vs. 77-100%, respectively). As such, predation seems to be the primary factor driving across-ecosystem distributions of this fish, and thus the primary reason why mangrove and seagrass habitats function as nursery habitat. Identifying the mechanisms that lead to such distributions is critical to develop appropriate conservation initiatives, identify essential fish habitat, and predict impacts associated with environmental change.
Figures




Similar articles
-
Effects of marine reserves versus nursery habitat availability on structure of reef fish communities.PLoS One. 2012;7(6):e36906. doi: 10.1371/journal.pone.0036906. Epub 2012 Jun 4. PLoS One. 2012. PMID: 22675474 Free PMC article.
-
The mangrove nursery paradigm revisited: otolith stable isotopes support nursery-to-reef movements by Indo-Pacific fishes.PLoS One. 2013 Jun 12;8(6):e66320. doi: 10.1371/journal.pone.0066320. Print 2013. PLoS One. 2013. PMID: 23776658 Free PMC article.
-
Distribution of fish in seagrass, mangroves and coral reefs: life-stage dependent habitat use in Honduras.Rev Biol Trop. 2012 Jun;60(2):683-98. doi: 10.15517/rbt.v60i2.3984. Rev Biol Trop. 2012. PMID: 23894938
-
The functional value of Caribbean coral reef, seagrass and mangrove habitats to ecosystem processes.Adv Mar Biol. 2006;50:57-189. doi: 10.1016/S0065-2881(05)50002-6. Adv Mar Biol. 2006. PMID: 16782451 Review.
-
[Metamorphosis of marine fish larvae and thyroid hormones].Biol Aujourdhui. 2019;213(1-2):27-33. doi: 10.1051/jbio/2019010. Epub 2019 Jul 5. Biol Aujourdhui. 2019. PMID: 31274100 Review. French.
Cited by
-
Predators and habitat association of post-settlement snapper (Chrysophrys auratus).J Fish Biol. 2022 Dec;101(6):1509-1521. doi: 10.1111/jfb.15222. Epub 2022 Oct 13. J Fish Biol. 2022. PMID: 36131511 Free PMC article.
-
Mangrove habitat use by juvenile reef fish: meta-analysis reveals that tidal regime matters more than biogeographic region.PLoS One. 2014 Dec 31;9(12):e114715. doi: 10.1371/journal.pone.0114715. eCollection 2014. PLoS One. 2014. PMID: 25551761 Free PMC article.
-
Cross-shelf habitat shifts by red snapper (Lutjanus campechanus) in the Gulf of Mexico.PLoS One. 2019 Mar 14;14(3):e0213506. doi: 10.1371/journal.pone.0213506. eCollection 2019. PLoS One. 2019. PMID: 30870449 Free PMC article.
-
Growth dynamics of juvenile loggerhead sea turtles undergoing an ontogenetic habitat shift.Oecologia. 2017 Apr;183(4):1087-1099. doi: 10.1007/s00442-017-3832-5. Epub 2017 Feb 16. Oecologia. 2017. PMID: 28210809
-
Effects of marine reserves versus nursery habitat availability on structure of reef fish communities.PLoS One. 2012;7(6):e36906. doi: 10.1371/journal.pone.0036906. Epub 2012 Jun 4. PLoS One. 2012. PMID: 22675474 Free PMC article.
References
-
- Adams AJ, Dahlgren CP, Kellison GT, Kendall MS, Layman CA, Ley JA, Nagelkerken I, Serafy JE. Nursery function of tropical back-reef systems. Mar Ecol Prog Ser. 2006;318:287–301. doi: 10.3354/meps318287. - DOI
-
- Aronson RB, Heck KL. Tethering experiments and hypothesis testing in ecology. Mar Ecol Prog Ser. 1995;121:307–309. doi: 10.3354/meps121307. - DOI
-
- Atkinson A, Siegel V, Pakhomov EA, Rothery P, Loeb V, Ross RM, Quetin LB, Schmidt K, Fretwell P, Murphy EJ, Tarling GA, Fleming AH. Oceanic circumpolar habitats of Antarctic krill. Mar Ecol Prog Ser. 2008;362:1–23. doi: 10.3354/meps07498. - DOI
-
- Baker R, Sheaves M. Shallow-water refuge paradigm: conflicting evidence from tethering experiments in a tropical estuary. Mar Ecol Prog Ser. 2007;349:13–22. doi: 10.3354/meps07102. - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
