

Dynamic origins of differential RNA binding function in two dsRBDs from the miRNA "microprocessor" complex
- PMID: 21073201
- PMCID: PMC3565223
- DOI: 10.1021/bi1015716
Dynamic origins of differential RNA binding function in two dsRBDs from the miRNA "microprocessor" complex
Abstract
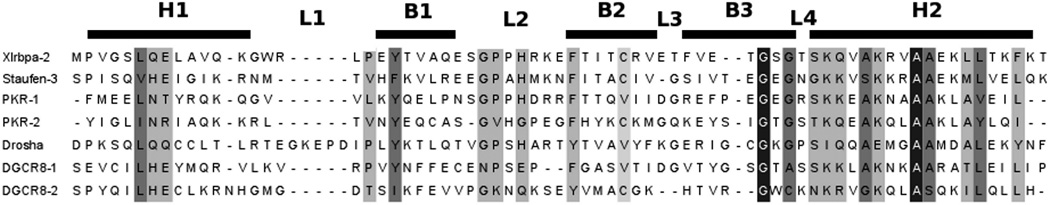
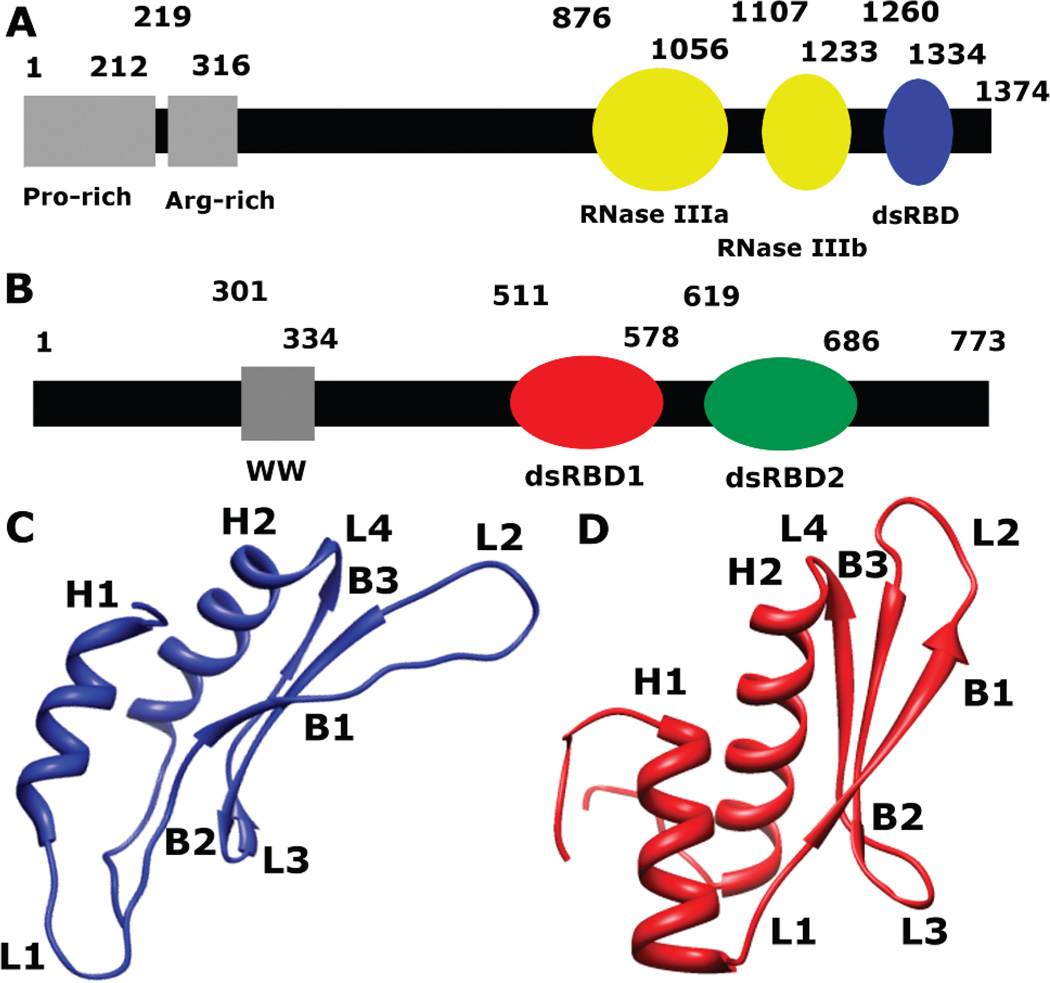
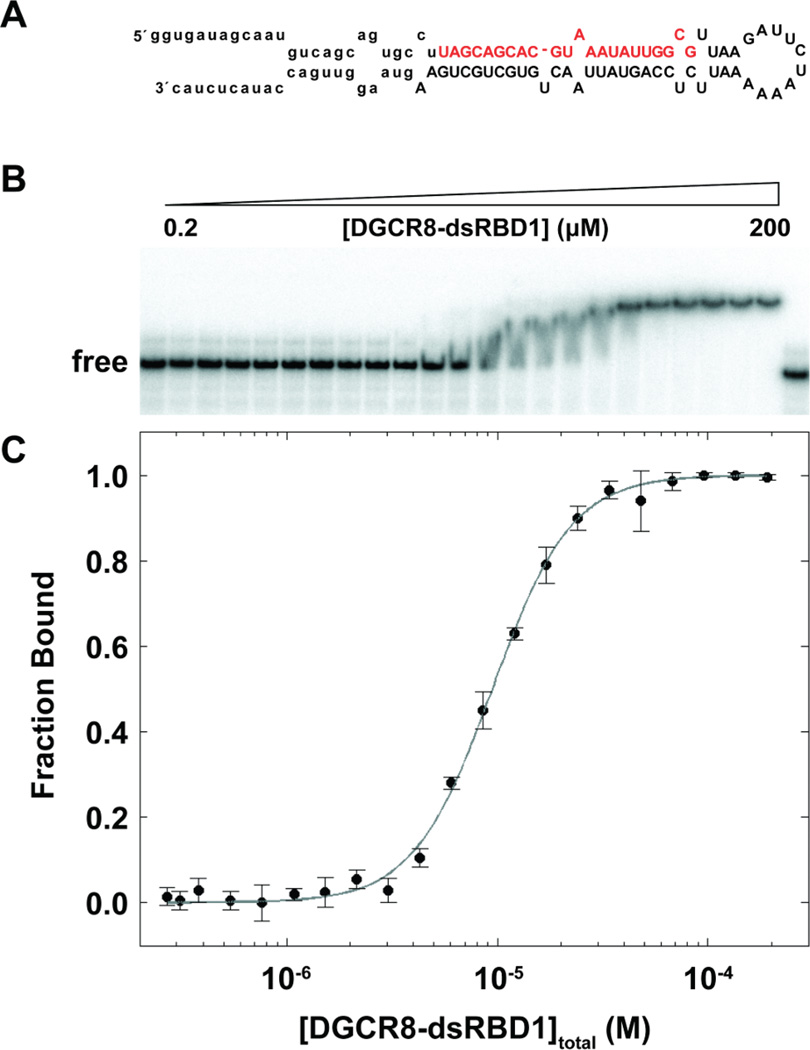
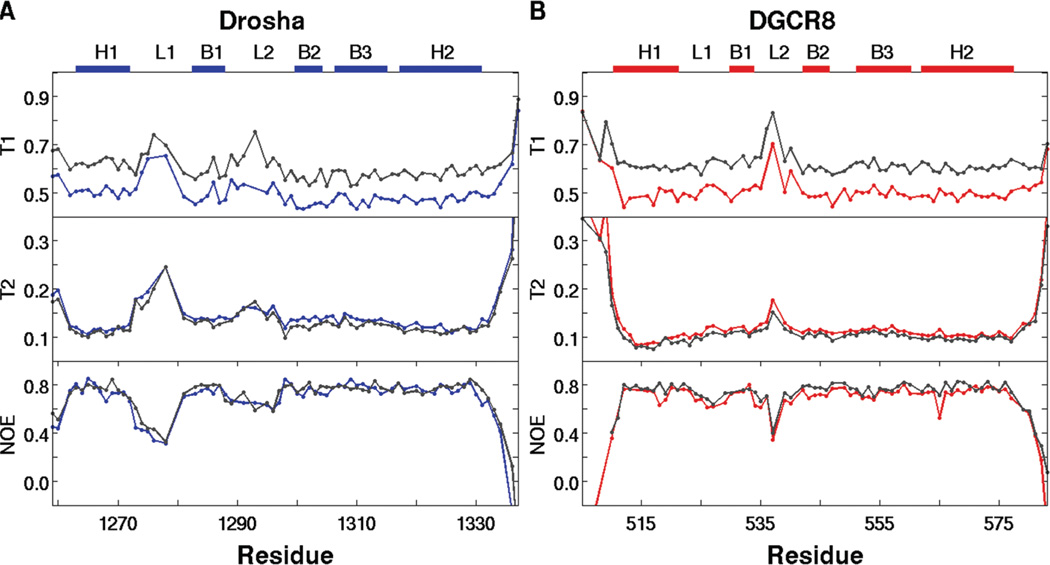
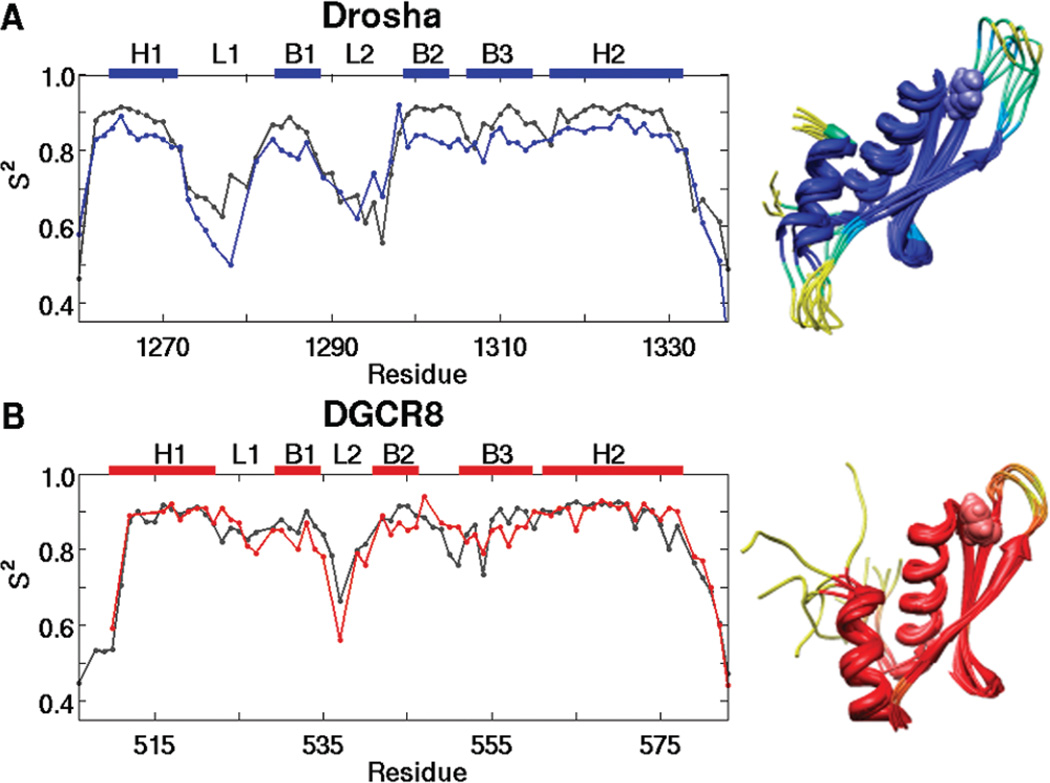
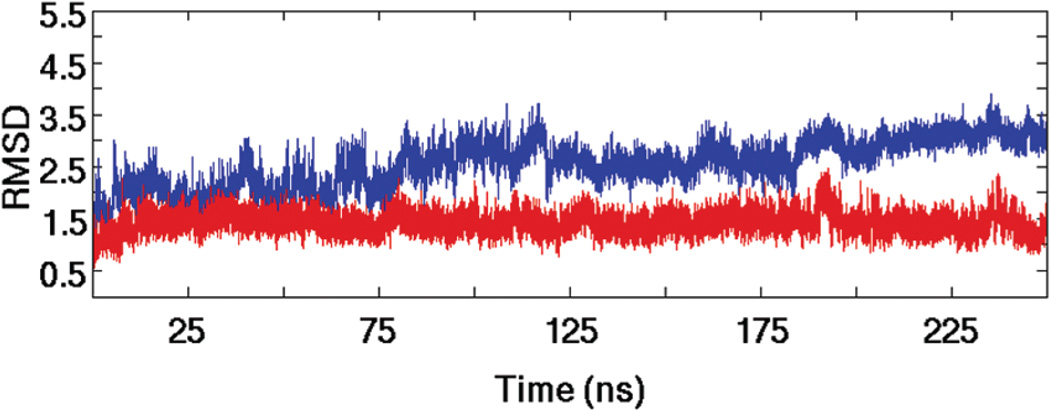
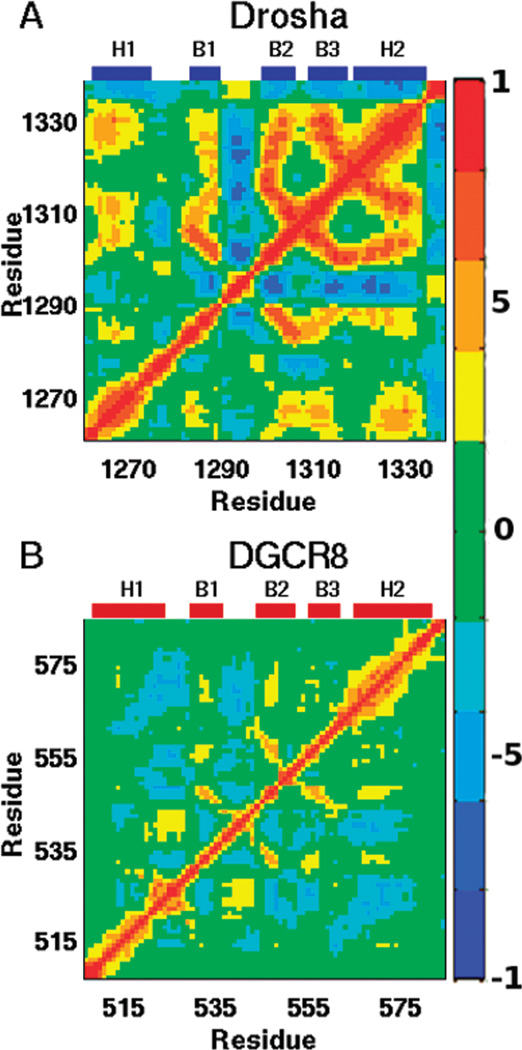
MicroRNAs (miRNAs) affect gene regulation by base pairing with mRNA and contribute to the control of cellular homeostasis. The first step in miRNA maturation is conducted in the nucleus by the "microprocessor" complex made up of an RNase III enzyme, Drosha, that contains one dsRNA binding domain (dsRBD), and DGCR8, that contains two dsRBDs in tandem. The crystal structure of DGCR8-Core (493-720), containing both dsRBDs, and the NMR solution structure of Drosha-dsRBD (1259-1337) have been reported, but the solution dynamics have not been explored for any of these dsRBDs. To better define the mechanism of dsRNA binding and thus the nuclear maturation step of miRNA processing, we report NMR spin relaxation and MD simulations of Drosha-dsRBD (1259-1337) and DGCR8-dsRBD1 (505-583). The study was motivated by electrophoretic mobility shift assays (EMSAs) of the two dsRBDs, which showed that Drosha-dsRBD does not bind a representative miRNA but isolated DGCR8-dsRBD1 does (K(d) = 9.4 ± 0.4 μM). Our results show that loop 2 in both dsRBDs is highly dynamic but the pattern of the correlations observed in MD is different for the two proteins. Additionally, the extended loop 1 of Drosha-dsRBD is more flexible than the corresponding loop in DGCR8-dsRBD1 but shows no correlation with loop 2, which potentially explains the lack of dsRNA binding by Drosha-dsRBD in the absence of the RNase III domains. The results presented in this study provide key structural and dynamic features of dsRBDs that contribute to the binding mechanism of these domains to dsRNA.
Figures
References
-
- Zhao Y, Srivastava D. A developmental view of micro-RNA function. Trends Biochem. Sci. 2007;32:189–197. - PubMed
-
- Wang Y, Liang Y, Lu Q. MicroRNA epigenetic alterations: predicting biomarkers and therapeutic targets in human diseases. Clin. Genet. 2008;74:307–315. - PubMed
-
- Negrini M, Ferracin M, Sabbioni S, Croce CM. MicroRNAs in human cancer: from research to therapy. J. Cell Sci. 2007;120:1833–1840. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
