

Review
doi: 10.1016/j.cell.2010.10.008.
Pluripotency and cellular reprogramming: facts, hypotheses, unresolved issues
Affiliations
- PMID: 21074044
- PMCID: PMC3032267
- DOI: 10.1016/j.cell.2010.10.008
Item in Clipboard
Review
Pluripotency and cellular reprogramming: facts, hypotheses, unresolved issues
Cell.
.
Abstract
Direct reprogramming of somatic cells to induced pluripotent stem cells by ectopic expression of defined transcription factors has raised fundamental questions regarding the epigenetic stability of the differentiated cell state. In addition, evidence has accumulated that distinct states of pluripotency can interconvert through the modulation of both cell-intrinsic and exogenous factors. To fully realize the potential of in vitro reprogrammed cells, we need to understand the molecular and epigenetic determinants that convert one cell type into another. Here we review recent advances in this rapidly moving field and emphasize unresolved and controversial questions.
Copyright © 2010 Elsevier Inc. All rights reserved.
Figures
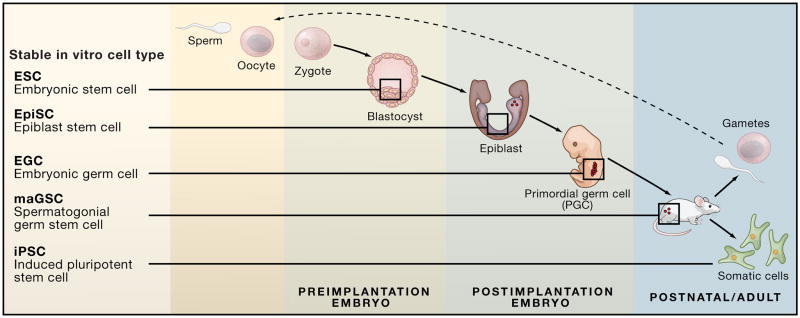
Different types of pluripotent cells can be derived by explanting cells at various stages of early embryonic development. Induced pluripotent stem cells (iPSCs) are derived by direct reprogramming of somatic cells in vitro.
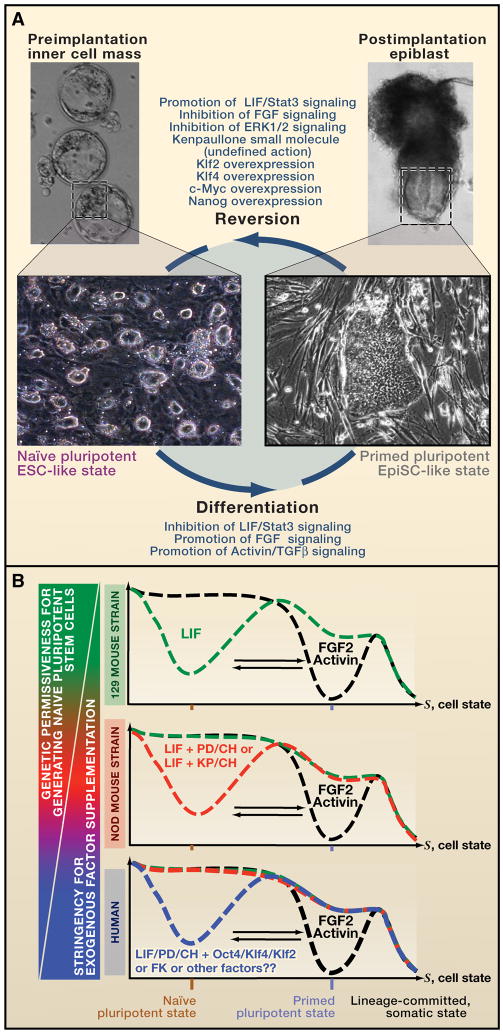
A) Naïve and primed cell types represent distinct states of gene expression states, corresponding to that observed in the pre-implantation inner cell mass and post-implantation epiblast, respectively. To stabilize the primed state in vitro, supplementation with basic fibroblast growth factor (bFGF) and Activin support the core transcriptional circuitry governing this state. In contrast, the naïve state has distinct active signaling pathways and thus requires different exogenous signals to induce and stabilize this state in vitro. Depending on the genetic background, combinations of these perturbations promote reversion to naïve pluripotency and differentiation into the primed pluripotent state. B) In this illustration, stabilization of the naïve and primed states in vitro by leukemia inhibitory factor (LIF) and bFGF/Activin signaling, respectively, is depicted as creating a well in a landscape. These factors can promote or antagonize inter-conversion between the states. LIF signaling promotes transfer of primed cells to the naïve state and continuously prevents differentiation of the naïve state. Shielding bFGF/Activin signaling can further enhance conversion into naïve cells, as this signaling pathway is inhibitory to the naïve state. In the non-permissive NOD mouse strain, LIF signaling alone is not sufficient to maintain the naïve state in vitro, as pluripotent cells derived from the inner cell mass or by in vitro reprogramming assume a primed state that can be stabilized by bFGF/Activin. However, the modulation of additional pathways, which are known to promote the naïve state and prevent differentiation, allowed derivation of naïve pluripotent cells from the NOD strain. This was achieved by altering the culture conditions, either with small molecules or by adding LIF, increasing Wnt signaling (small molecule ‘CH’), and inhibiting ERK1/2 (small molecule ‘PD’). The human genetic background is less permissive, as it required modulation of additional signaling pathways to induce the naïve state.
exogenous factors, genetic background, and epigenetics of the tissue origin of cells.
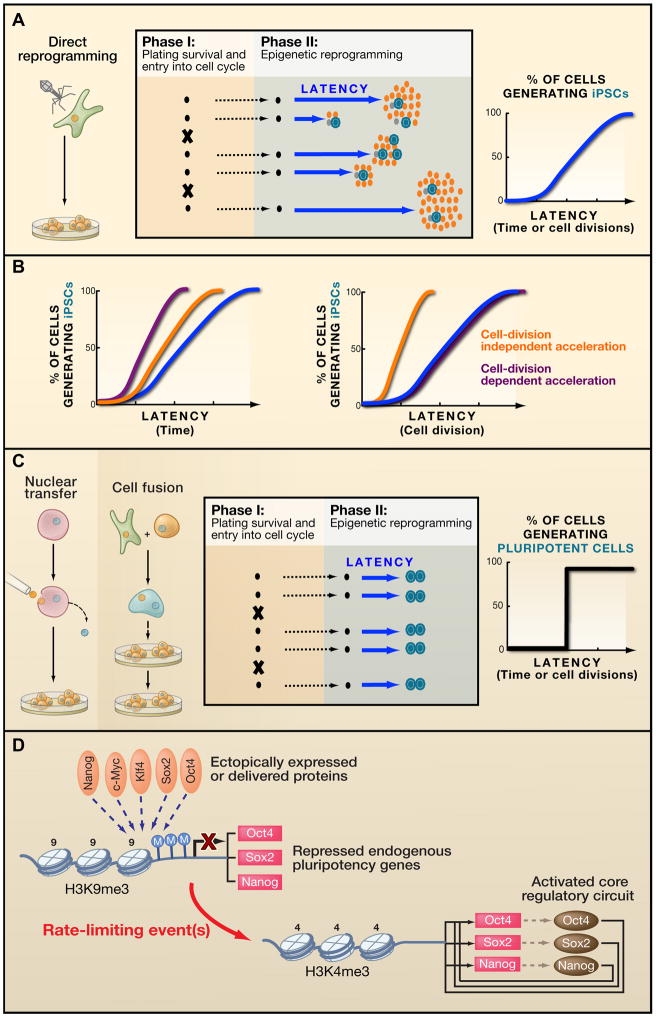
A) In direct reprogramming, the progression of clonal populations to a reprogrammed state first involves an initial technical phase (I), which depends on the survival of plated cells and their entry into the cell cycle. Once cell division occurs, most critical phase begins. During this second phase (II), direct reprogramming involves a stochastic event because clonal populations do not give rise to iPSCs at the same time after Phase I. This variation in latency is represented by the blue line. B) Phase II can be accelerated by two mechanisms involving cell division (purple) or mechanisms independent of cell division (orange). C) Like with direct reprogramming, the progression of reprogramming by nuclear transfer and cell fusion involves two phases. However, compared to direct reprogramming, , much less heterogeneity is observed nuclear transfer and cell fusion. For one, partially reprogrammed lines are not observed with nuclear transfer or fusion, and the reprogramming is hypothesized to progress in a more deterministic manner. This suggests that the current protocols of direct reprogramming are not optimal and may be accelerated by the supplementing with more factors to eliminate the stochasticity and achieve the deterministic conversion observed in fusion and nuclear transfer if. D) One key rate-limiting step during direct reprogramming may be reactivation of the core pluripotency regulatory circuitry.
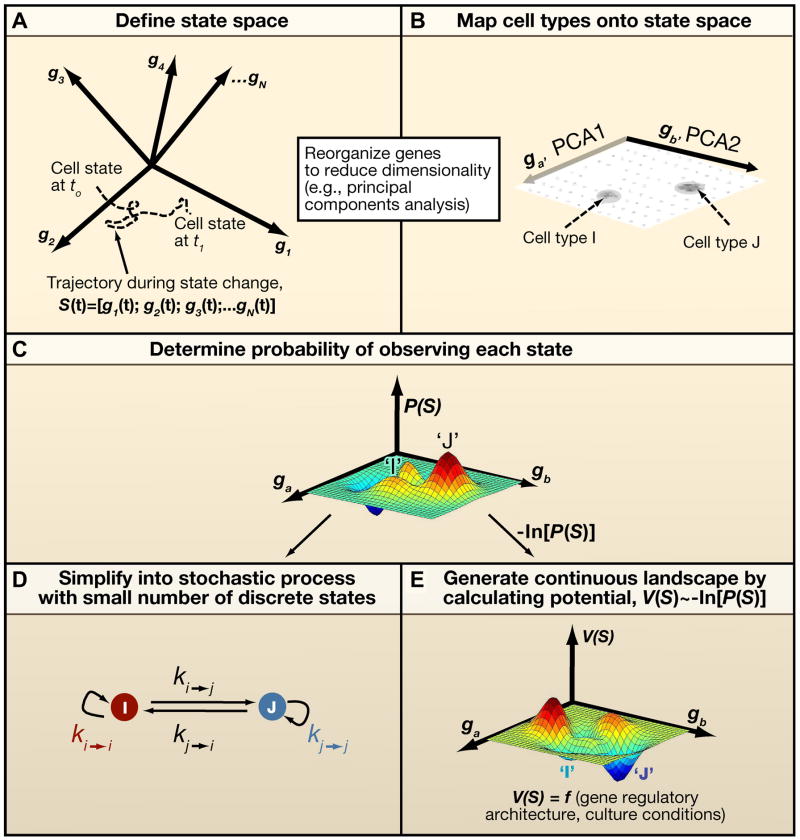
A) First, a set of characteristics must be identified as being informative of the for the transition between two cell states of interest. These characteristics usually consist of levels of gene expression or levels of epigenetic methylation or acetylation marks on DNA or chromatin. For simplicity, we show here a state space generated by the level of N different genes, g1 to gN, where each arrow represents an axis corresponding to the expression level of that particular gene transcript (N~104 for mammalian cells). A cell at any time t exists in a point in this space, and its state can change with time as a result of noise, reprogramming, or differentiation. One trajectory is plotted, and the vector Ŝ, which consists of gene expression levels changing with time, fully describes the cell state transition during this time. B) N can be reduced to a more manageable 2–3 dimensions through statistical techniques, such as principal component analysis (PCA). Shown here is a two-dimensional representation of the state space in part A), where each axis (ga and gb from PCA) is a linear combination of particular genes. Stable cells in vitro exist at particular points on this graph. By mapping quantitatively the gene expression levels of several single stable cells in the same space, regions with high densities of spots define observable cell types in vitro (as type “I” and “J”). C) When a large sample of single cells is mapped in part B, the probability of occupying each point in this space can be calculated and plotted in a continuous fashion. D) The continuous probabilistic description of cell state in C can be simplified into a discrete representation, with a small number of discrete states that transition at particular rates, k. These rates, k, represent an average of all possible trajectories from region i to region j. E) Alternatively, a continuous description of the probabilities of staying at a particular point in state space can be represented as a landscape, by calculating −ln[P(Ŝ)] from part C at each point. This landscape, V(Ŝ), represents ‘energy barriers’ between transitions involving any two states and thus may provide a more thorough description of transitions than the description in D) .
References
-
- Bloom JD, Meyer MM, Meinhold P, Otey CR, MacMillan D, Arnold FH. Evolving strategies for enzyme engineering. Curr Opin Struct Biol. 2005;15:447–452. - PubMed
-
- Boland MJ, Hazen JL, Nazor KL, Rodriguez AR, Gifford W, Martin G, Kupriyanov S, Baldwin KK. Adult mice generated from induced pluripotent stem cells. Nature. 2009;461:91–94. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
