

Pausing of RNA polymerase II disrupts DNA-specified nucleosome organization to enable precise gene regulation
- PMID: 21074046
- PMCID: PMC2991113
- DOI: 10.1016/j.cell.2010.10.004
Pausing of RNA polymerase II disrupts DNA-specified nucleosome organization to enable precise gene regulation
Abstract
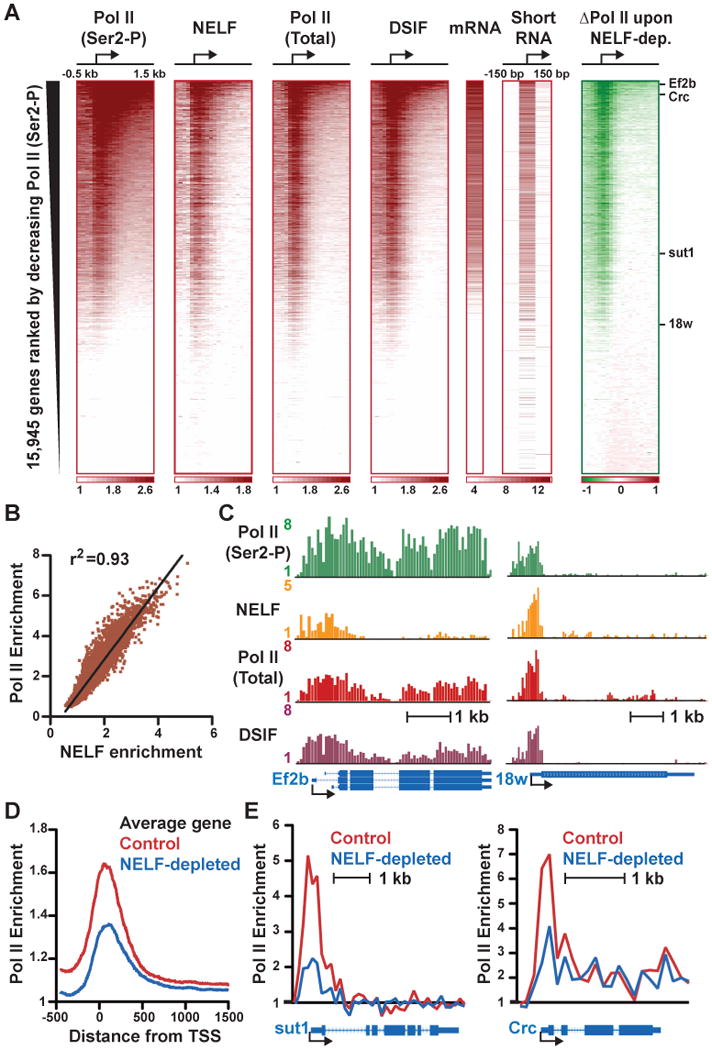
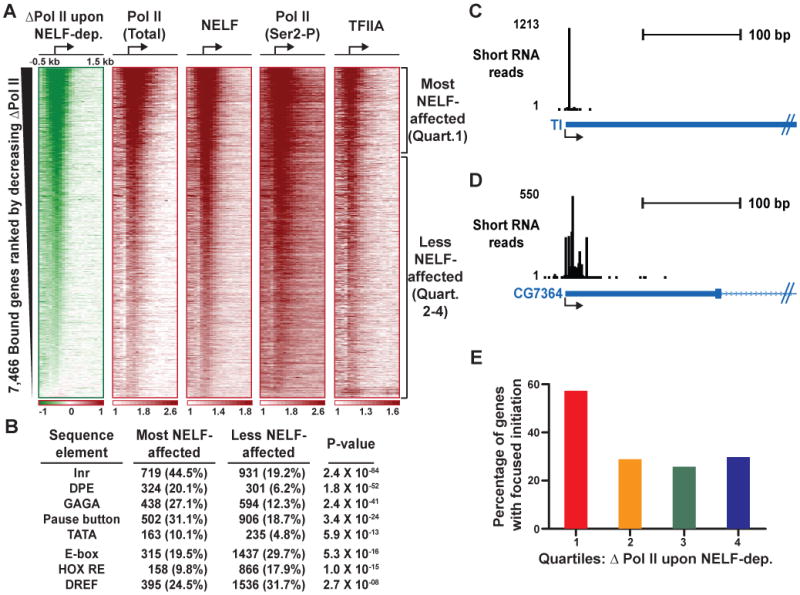
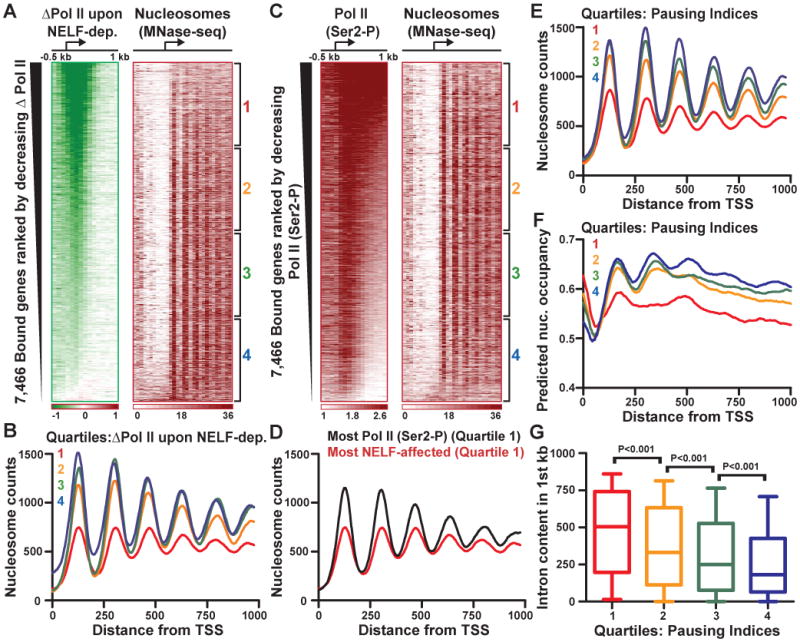
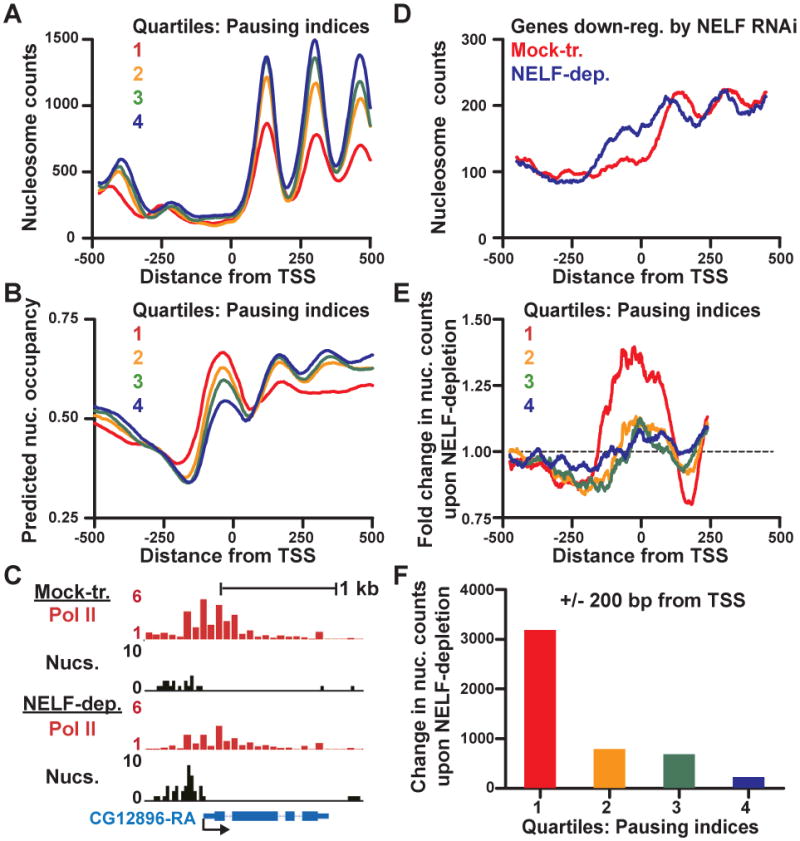
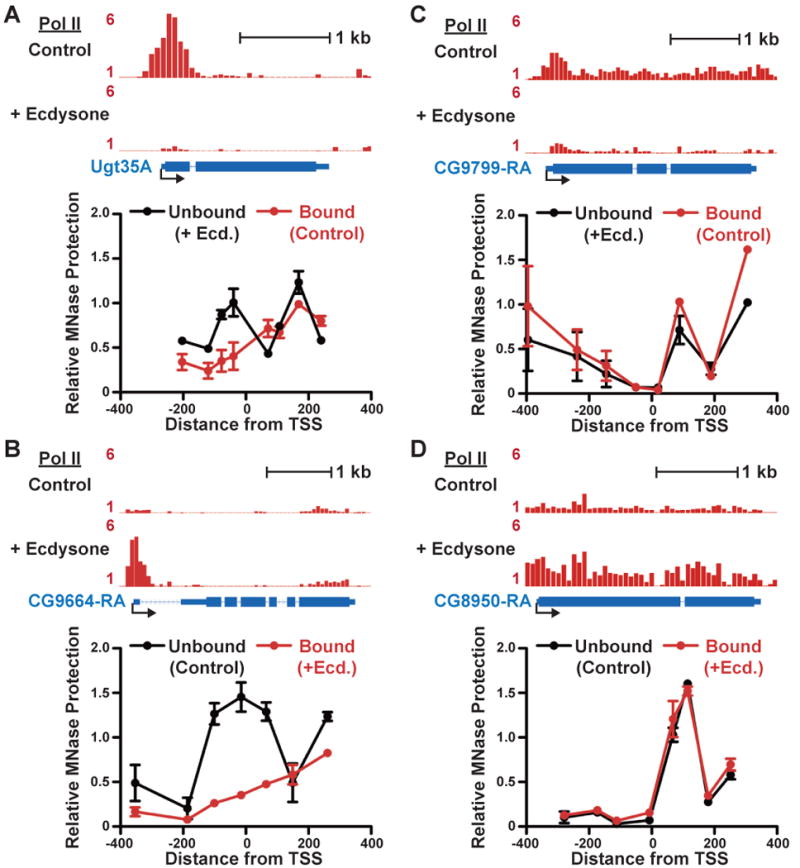
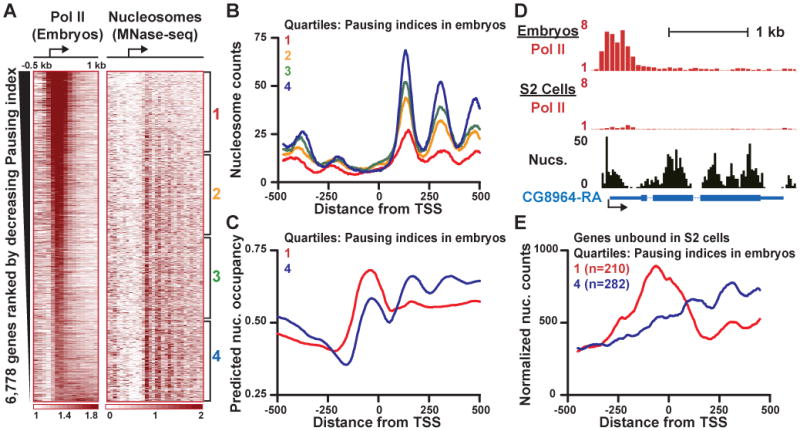
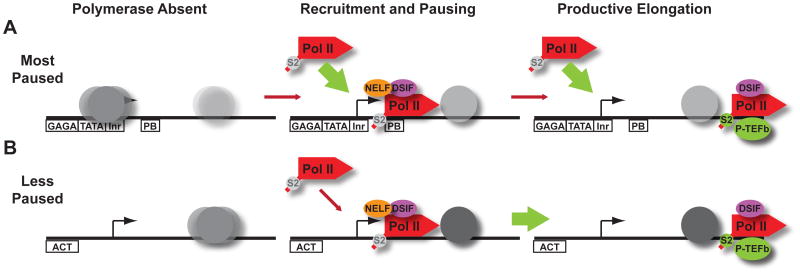
Metazoan transcription is controlled through either coordinated recruitment of transcription machinery to the gene promoter or regulated pausing of RNA polymerase II (Pol II) in early elongation. We report that a striking difference between genes that use these distinct regulatory strategies lies in the "default" chromatin architecture specified by their DNA sequences. Pol II pausing is prominent at highly regulated genes whose sequences inherently disfavor nucleosome formation within the gene but favor occlusion of the promoter by nucleosomes. In contrast, housekeeping genes that lack pronounced Pol II pausing show higher nucleosome occupancy downstream, but their promoters are deprived of nucleosomes regardless of polymerase binding. Our results indicate that a key role of paused Pol II is to compete with nucleosomes for occupancy of highly regulated promoters, thereby preventing the formation of repressive chromatin architecture to facilitate further or future gene activation.
Copyright © 2010 Elsevier Inc. All rights reserved.
Figures
Comment in
-
The meaning of pausing.Mol Cell. 2010 Nov 24;40(4):507-8. doi: 10.1016/j.molcel.2010.11.025. Mol Cell. 2010. PMID: 21095581
References
-
- Albert I, Mavrich TN, Tomsho LP, Qi J, Zanton SJ, Schuster SC, Pugh BF. Translational and rotational settings of H2A.Z nucleosomes across the Saccharomyces cerevisiae genome. Nature. 2007;446:572–576. - PubMed
-
- Basehoar AD, Zanton SJ, Pugh BF. Identification and distinct regulation of yeast TATA box-containing genes. Cell. 2004;116:699–709. - PubMed
-
- Cairns BR. The logic of chromatin architecture and remodelling at promoters. Nature. 2009;461:193–198. - PubMed
-
- Cheng B, Price DH. Properties of RNA Polymerase II Elongation Complexes Before and After the P-TEFb-mediated Transition into Productive Elongation. The Journal of biological chemistry. 2007;282:21901–21912. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
