

A genome-wide Drosophila screen for heat nociception identifies α2δ3 as an evolutionarily conserved pain gene
- PMID: 21074052
- PMCID: PMC3040441
- DOI: 10.1016/j.cell.2010.09.047
A genome-wide Drosophila screen for heat nociception identifies α2δ3 as an evolutionarily conserved pain gene
Abstract
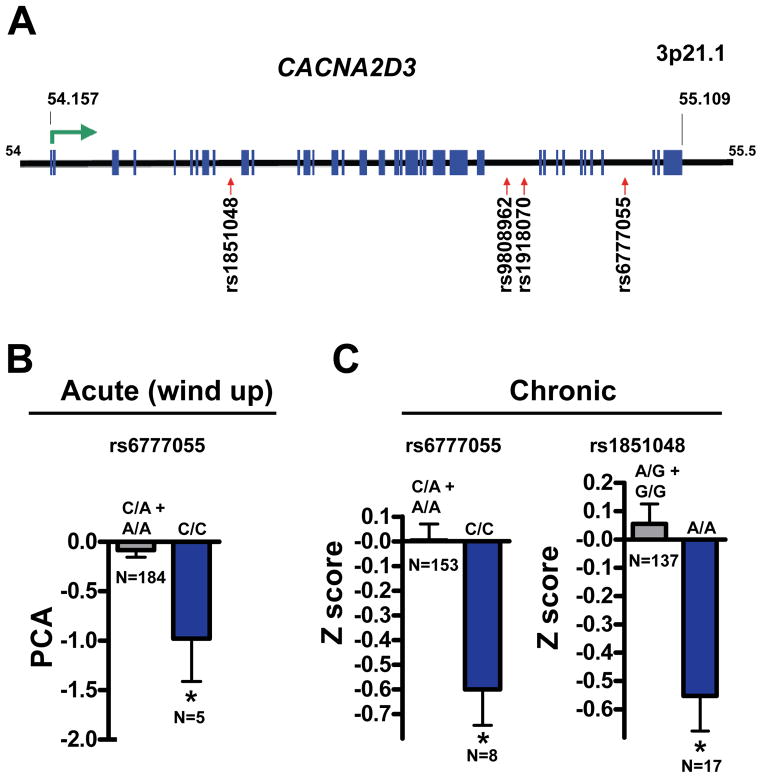
Worldwide, acute, and chronic pain affects 20% of the adult population and represents an enormous financial and emotional burden. Using genome-wide neuronal-specific RNAi knockdown in Drosophila, we report a global screen for an innate behavior and identify hundreds of genes implicated in heat nociception, including the α2δ family calcium channel subunit straightjacket (stj). Mice mutant for the stj ortholog CACNA2D3 (α2δ3) also exhibit impaired behavioral heat pain sensitivity. In addition, in humans, α2δ3 SNP variants associate with reduced sensitivity to acute noxious heat and chronic back pain. Functional imaging in α2δ3 mutant mice revealed impaired transmission of thermal pain-evoked signals from the thalamus to higher-order pain centers. Intriguingly, in α2δ3 mutant mice, thermal pain and tactile stimulation triggered strong cross-activation, or synesthesia, of brain regions involved in vision, olfaction, and hearing.
Copyright © 2010 Elsevier Inc. All rights reserved.
Figures






Comment in
-
A straightjacket for pain?Cell. 2010 Nov 12;143(4):505-7. doi: 10.1016/j.cell.2010.10.038. Cell. 2010. PMID: 21074043
References
-
- Aleman A, Rutten GJ, Sitskoorn MM, Dautzenberg G, Ramsey NF. Activation of striate cortex in the absence of visual stimulation: an fMRI study of synesthesia. Neuroreport. 2001;12:2827–2830. - PubMed
-
- Arthurs OJ, Boniface S. How well do we understand the neural origins of the fMRI BOLD signal? Trends in neurosciences. 2002;25:27–31. - PubMed
-
- Atlas SJ, Keller RB, Chang Y, Deyo RA, Singer DE. Surgical and nonsurgical management of sciatica secondary to a lumbar disc herniation: five-year outcomes from the Maine Lumbar Spine Study. Spine. 2001;26:1179–1187. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
