

Drosophila orthologue of WWOX, the chromosomal fragile site FRA16D tumour suppressor gene, functions in aerobic metabolism and regulates reactive oxygen species
- PMID: 21075834
- PMCID: PMC3016910
- DOI: 10.1093/hmg/ddq495
Drosophila orthologue of WWOX, the chromosomal fragile site FRA16D tumour suppressor gene, functions in aerobic metabolism and regulates reactive oxygen species
Abstract
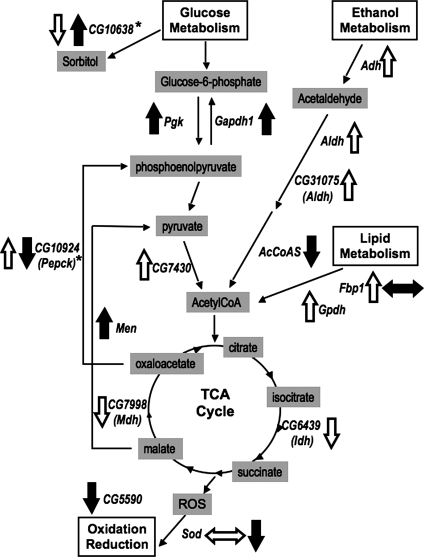
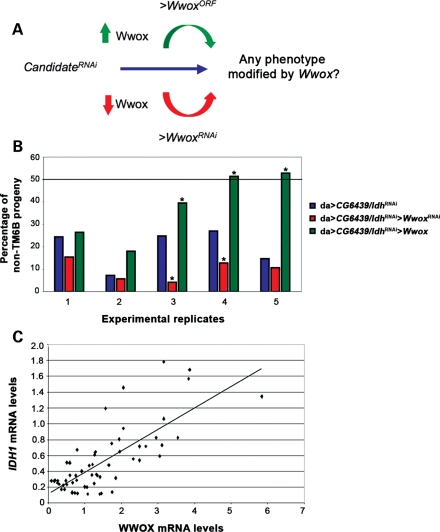
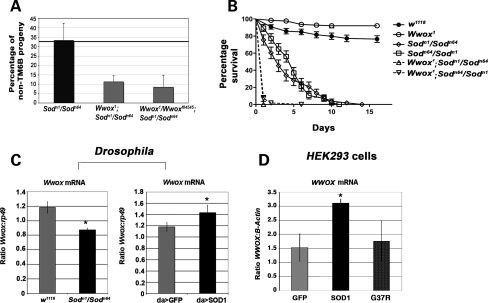
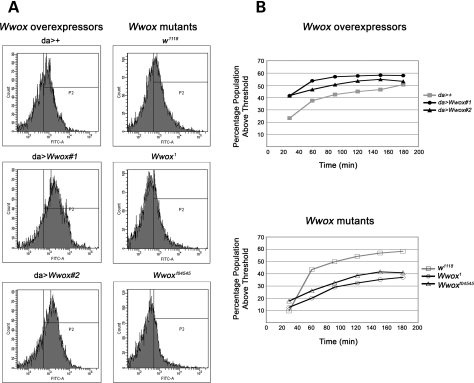
Common chromosomal fragile sites FRA3B and FRA16D are frequent sites of DNA instability in cancer, but their contribution to cancer cell biology is not yet understood. Genes that span these sites (FHIT and WWOX, respectively) are often perturbed (either increased or decreased) in cancer cells and both are able to suppress tumour growth. While WWOX has some tumour suppressor characteristics, its normal role and functional contribution to cancer has not been fully determined. We find that a significant proportion of Drosophila Wwox interactors identified by proteomics and microarray analyses have roles in aerobic metabolism. Functional relationships between Wwox and either CG6439/isocitrate dehydrogenase (Idh) or Cu-Zn superoxide dismutase (Sod) were confirmed by genetic interactions. In addition, altered levels of Wwox resulted in altered levels of endogenous reactive oxygen species. Wwox (like FHIT) contributes to pathways involving aerobic metabolism and oxidative stress, providing an explanation for the 'non-classical tumour suppressor' behaviour of WWOX. Fragile sites, and the genes that span them, are therefore part of a protective response mechanism to oxidative stress and likely contributors to the differences seen in aerobic glycolysis (Warburg effect) in cancer cells.
Figures

References
-
- Yunis J.J., Soreng A.L. Constitutive fragile sites and cancer. Science. 1984;226:1199–1204. - PubMed
-
- Richards R.I. Fragile and unstable chromosomes in cancer: causes and consequences. Trends Genet. 2001;17:339–345. - PubMed
-
- Stein C.K., Glover T.W., Palmer J.L., Glisson B.S. Direct correlation between FRA3B expression and cigarette smoking. Genes Chromosomes Cancer. 2002;34:333–340. - PubMed
-
- Glover T.W., Arlt M.F., Casper A.M., Durkin S.G. Mechanisms of common fragile site instability. Hum. Mol. Genet. 2005;14:R197–R205. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
