

Different contributions of HtrA protease and chaperone activities to Campylobacter jejuni stress tolerance and physiology
- PMID: 21075890
- PMCID: PMC3019702
- DOI: 10.1128/AEM.01603-10
Different contributions of HtrA protease and chaperone activities to Campylobacter jejuni stress tolerance and physiology
Abstract
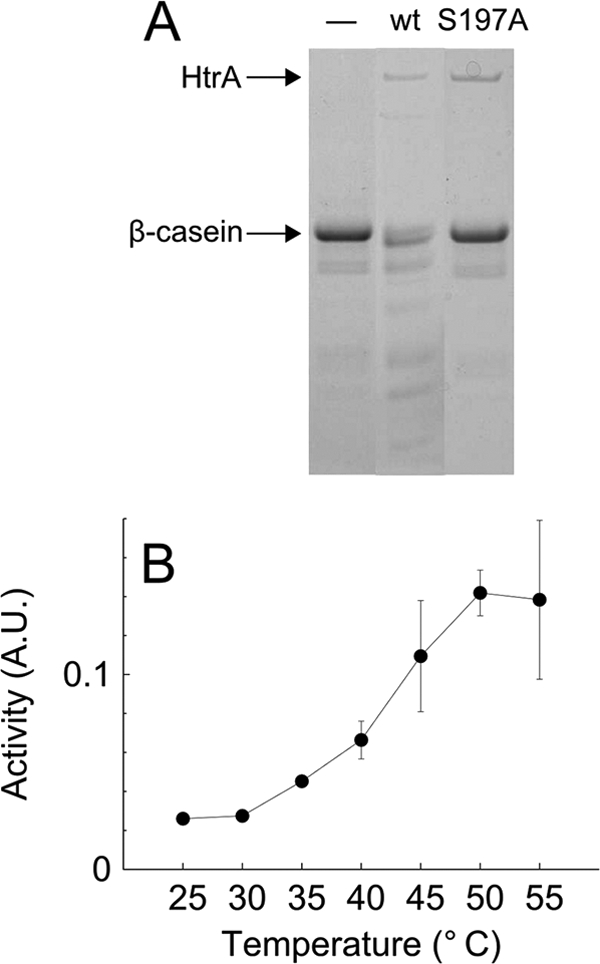
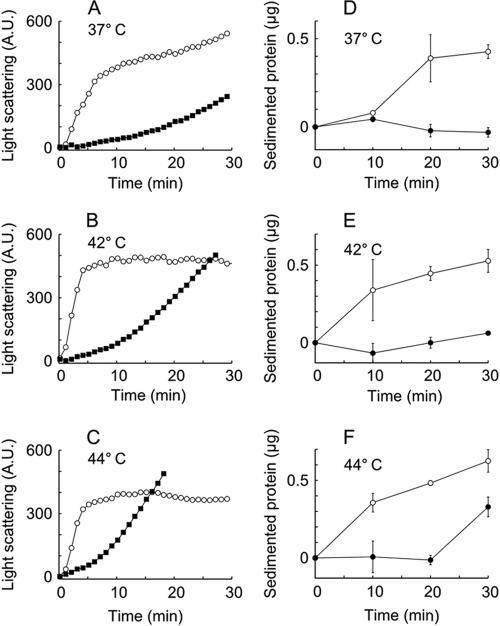
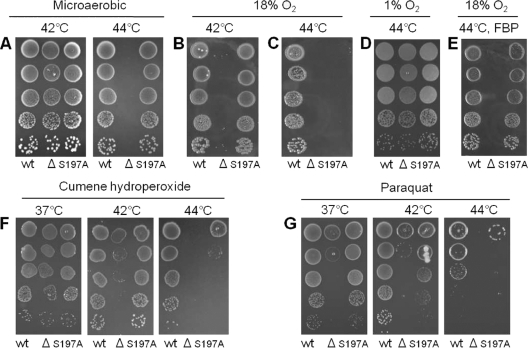
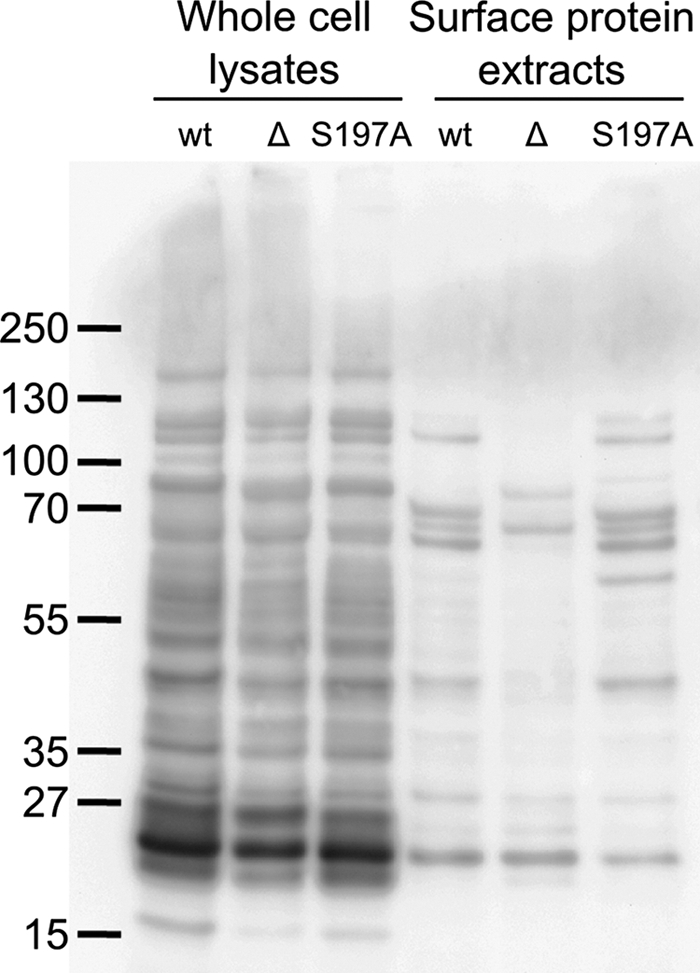
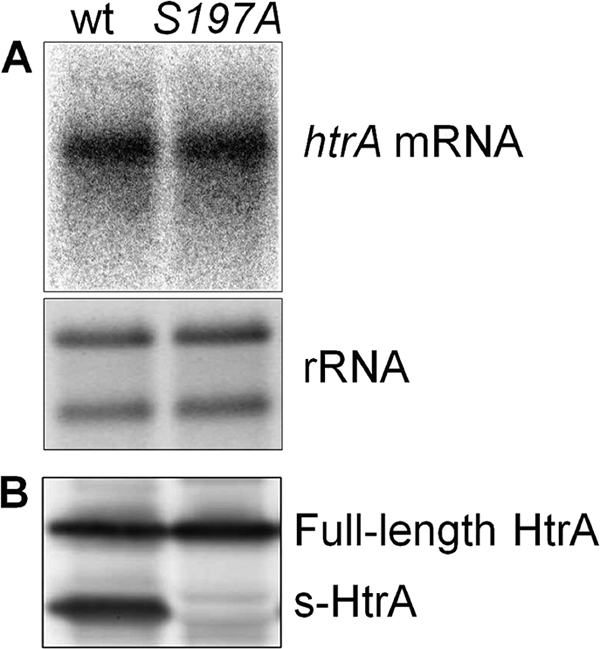
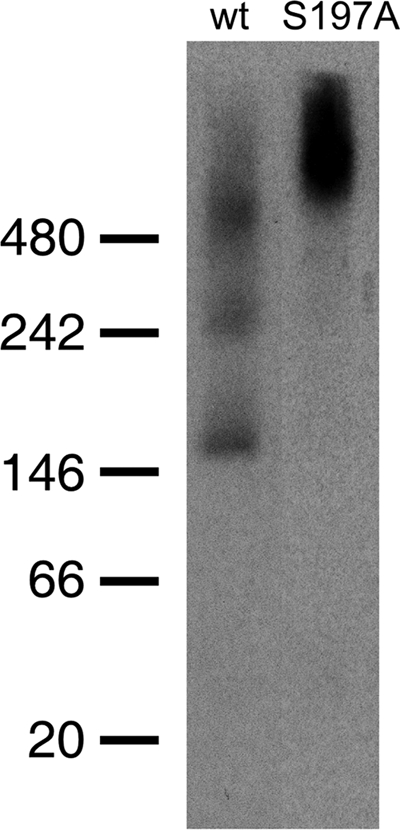
The microaerophilic bacterium Campylobacter jejuni is the most common cause of bacterial food-borne infections in the developed world. Tolerance to environmental stress relies on proteases and chaperones in the cell envelope, such as HtrA and SurA. HtrA displays both chaperone and protease activities, but little is known about how each of these activities contributes to stress tolerance in bacteria. In vitro experiments showed temperature-dependent protease and chaperone activities of C. jejuni HtrA. A C. jejuni mutant lacking only the protease activity of HtrA was used to show that the HtrA chaperone activity is sufficient for growth at high temperature or under oxidative stress, whereas the HtrA protease activity is essential only under conditions close to the growth limit for C. jejuni. However, the protease activity was required to prevent induction of the cytoplasmic heat shock response even under optimal growth conditions. Interestingly, the requirement of HtrA at high temperatures was found to depend on the oxygen level, and our data suggest that HtrA may protect oxidatively damaged proteins. Finally, protease activity stimulates HtrA production and oligomer formation, suggesting that a regulatory role depends on the protease activity of HtrA. Studying a microaerophilic organism encoding only two known periplasmic chaperones (HtrA and SurA) revealed an efficient HtrA chaperone activity and proposed multiple roles of the protease activity, increasing our understanding of HtrA in bacterial physiology.
Figures

References
-
- Allos, B. M. 2001. Campylobacter jejuni infections: update on emerging issues and trends. Clin. Infect. Dis. 32:1201-1206. - PubMed
-
- Arie, J. P., N. Sassoon, and J. M. Betton. 2001. Chaperone function of FkpA, a heat shock prolyl isomerase, in the periplasm of Escherichia coli. Mol. Microbiol. 39:199-210. - PubMed
-
- Asakura, H., M. Yamasaki, S. Yamamoto, and S. Igimi. 2007. Deletion of peb4 gene impairs cell adhesion and biofilm formation in Campylobacter jejuni. FEMS Microbiol. Lett. 275:278-285. - PubMed
-
- Atack, J. M., and D. J. Kelly. 2009. Oxidative stress in Campylobacter jejuni: responses, resistance and regulation. Future Microbiol. 4:677-690. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
