

Zinc requirement during meiosis I-meiosis II transition in mouse oocytes is independent of the MOS-MAPK pathway
- PMID: 21076080
- PMCID: PMC3043131
- DOI: 10.1095/biolreprod.110.086488
Zinc requirement during meiosis I-meiosis II transition in mouse oocytes is independent of the MOS-MAPK pathway
Abstract
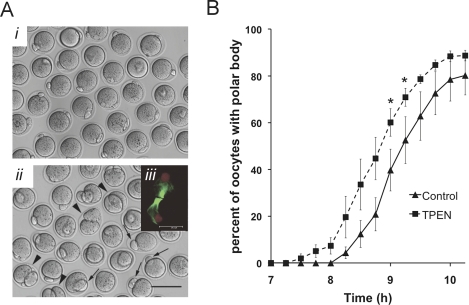
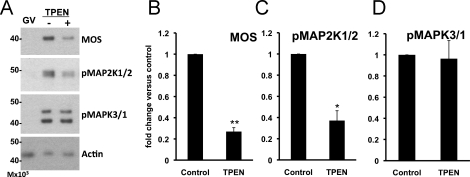
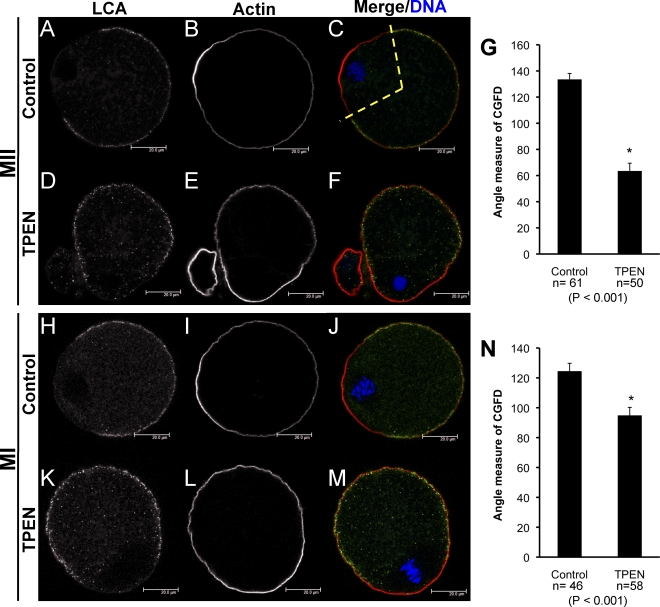
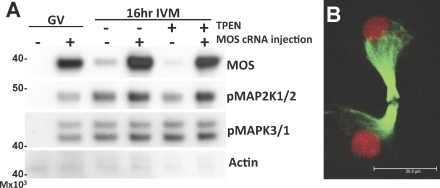
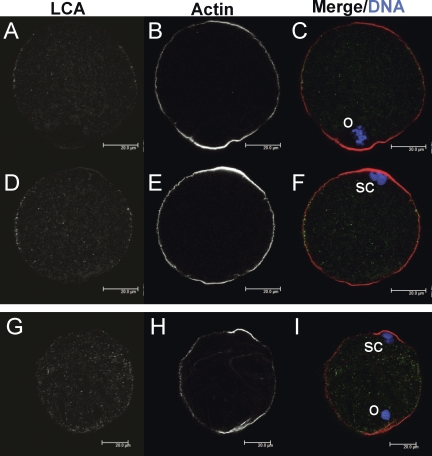
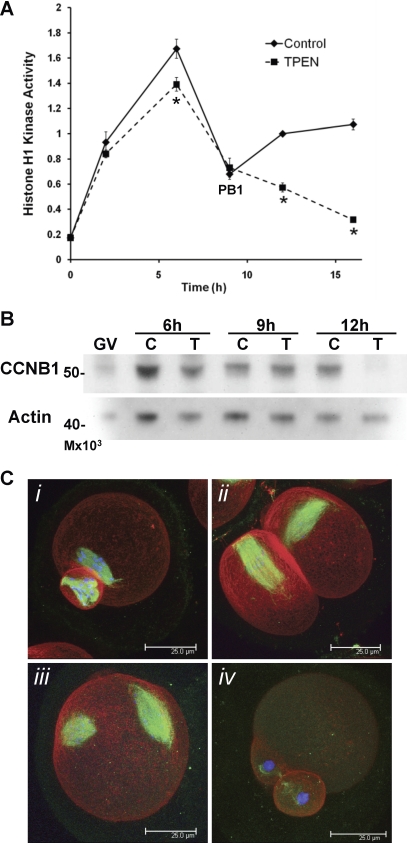
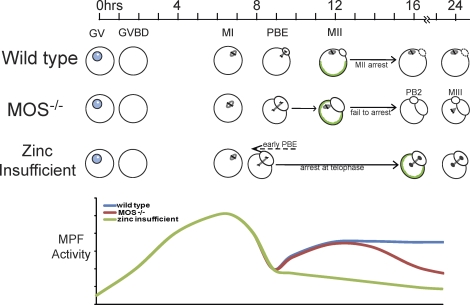
Zinc is essential for many biological processes, including proper functioning of gametes. We recently reported that zinc levels rise by over 50% during oocyte maturation and that attenuation of zinc availability during this period could be achieved using the membrane-permeable heavy metal chelator N,N,N',N'-tetrakis(2-pyridylmethyl)ethylenediamine (TPEN). This zinc insufficiency resulted in formation of large polar bodies, failure to establish metaphase II arrest, and impaired establishment of cortical polarity. As these phenotypes resemble those of MOS null oocytes, we examined the impact of zinc insufficiency on the MOS-MAPK pathway. Reduced levels of both MOS protein and phosphorylation of MAP2K1/2 are observed in zinc-insufficient oocytes; however, these differences appear only after completion of the first meiotic division. In addition, activation of the downstream effector of the MOS pathway, MAPK3/1, is not affected by zinc insufficiency, and reduced MOS levels are observed only with the presence of TPEN after the first polar body extrusion. These data are inconsistent with the hypothesis that reduced MOS mediates the observed phenotype. Finally, MOS overexpression does not rescue the phenotype of zinc-insufficient oocytes, confirming that the observed disruption of asymmetric division and spindle abnormalities cannot be attributed to impaired MOS signaling. Zinc-insufficient oocytes do not increase maturation promoting factor (MPF) activity following the first meiotic division, and increasing MPF activity through expression of nondegradable cyclin B1 partially rescues the ability of zinc-insufficient oocytes to enter metaphase II. Although we have shown that zinc has a novel role in the meiotic cell cycle, it is not mediated through the MOS-MAPK pathway.
Figures

References
-
- Vallee BL, Falchuk KH. The biochemical basis of zinc physiology. Physiol Rev 1993; 73: 79 118 - PubMed
-
- Andrews JC, Nolan JP, Hammerstedt RH, Bavister BD. Role of zinc during hamster sperm capacitation. Biol Reprod 1994; 51: 1238 1247 - PubMed
-
- de Lamirande E, Lamothe G, Villemure M. Control of superoxide and nitric oxide formation during human sperm capacitation. Free Radic Biol Med 2009; 46: 1420 1427 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
