

Importance of the proline-rich multimerization domain on the oligomerization and nucleic acid binding properties of HIV-1 Vif
- PMID: 21076154
- PMCID: PMC3064812
- DOI: 10.1093/nar/gkq979
Importance of the proline-rich multimerization domain on the oligomerization and nucleic acid binding properties of HIV-1 Vif
Abstract
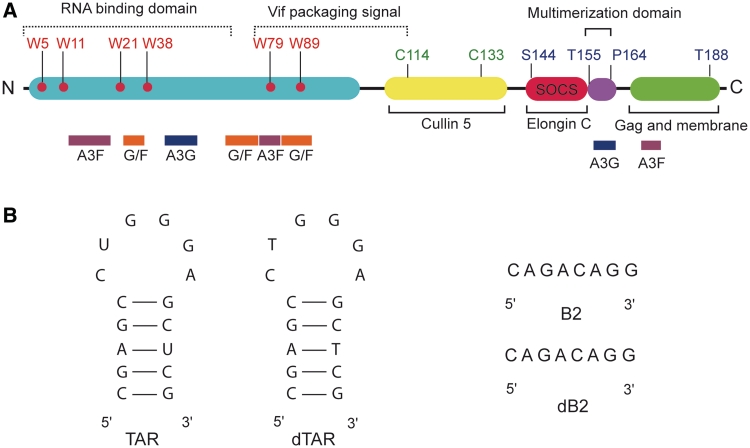
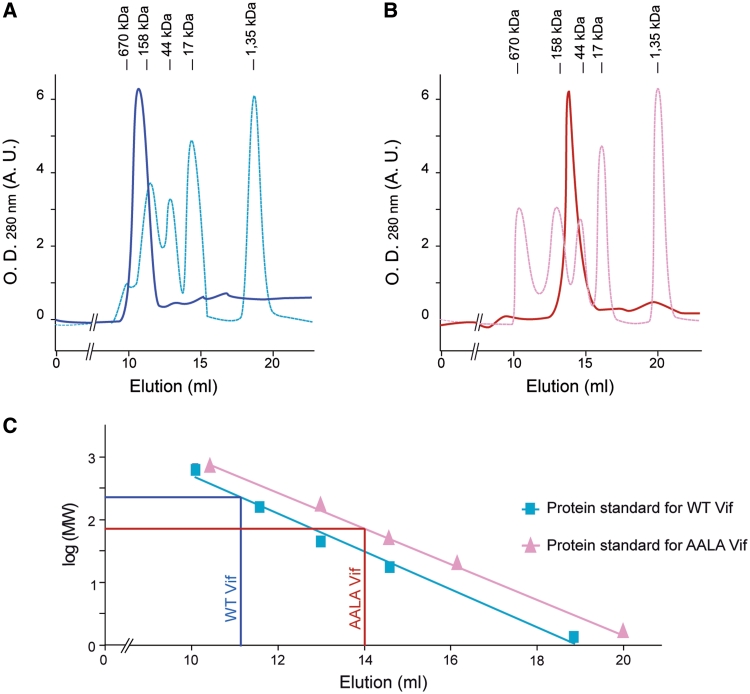
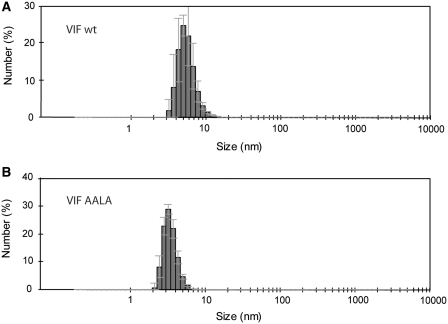
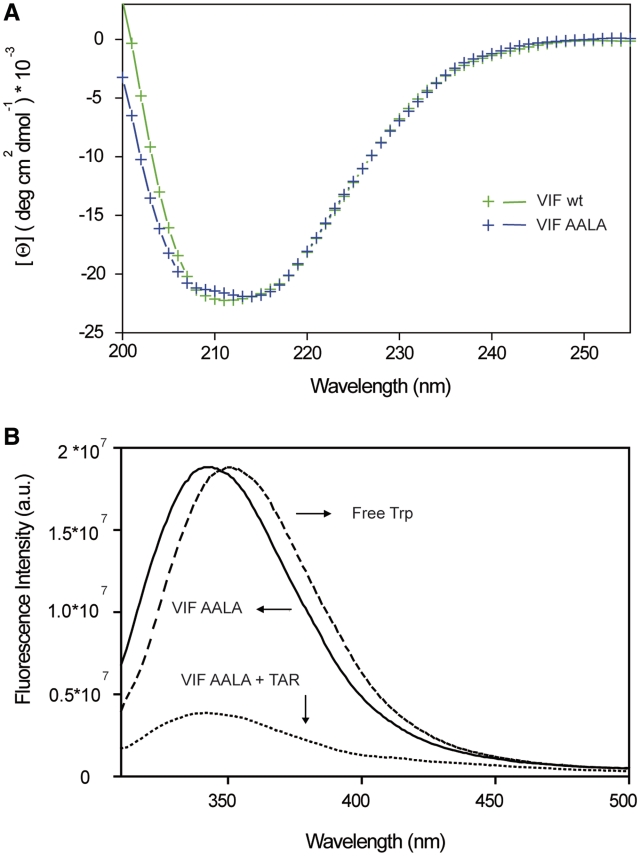
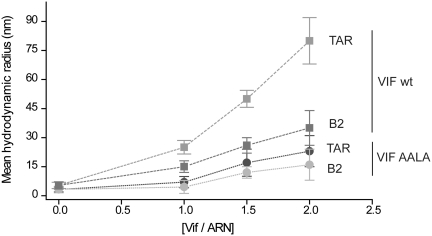
The HIV-1 viral infectivity factor (Vif) is required for productive infection of non-permissive cells, including most natural HIV-1 targets, where it counteracts the antiviral activities of the cellular cytosine deaminases APOBEC-3G (A3G) and A3F. Vif is a multimeric protein and the conserved proline-rich domain (161)PPLP(164) regulating Vif oligomerization is crucial for its function and viral infectivity. Here, we expressed and purified wild-type Vif and a mutant protein in which alanines were substituted for the proline residues of the (161)PPLP(164) domain. Using dynamic light scattering, circular dichroism and fluorescence spectroscopy, we established the impact of these mutations on Vif oligomerization, secondary structure content and nucleic acids binding properties. In vitro, wild-type Vif formed oligomers of five to nine proteins, while Vif AALA formed dimers and/or trimers. Up to 40% of the unbound wild-type Vif protein appeared to be unfolded, but binding to the HIV-1 TAR apical loop promoted formation of β-sheets. Interestingly, alanine substitutions did not significantly affect the secondary structure of Vif, but they diminished its binding affinity and specificity for nucleic acids. Dynamic light scattering showed that Vif oligomerization, and interaction with folding-promoting nucleic acids, favor formation of high molecular mass complexes. These properties could be important for Vif functions involving RNAs.
Figures
Similar articles
-
APOBEC3G impairs the multimerization of the HIV-1 Vif protein in living cells.J Virol. 2013 Jun;87(11):6492-506. doi: 10.1128/JVI.03494-12. Epub 2013 Apr 10. J Virol. 2013. PMID: 23576497 Free PMC article.
-
The unique structure of the highly conserved PPLP region in HIV-1 Vif is critical for the formation of APOBEC3 recognition interfaces.mBio. 2025 Mar 12;16(3):e0333224. doi: 10.1128/mbio.03332-24. Epub 2025 Jan 21. mBio. 2025. PMID: 39835817 Free PMC article.
-
Characterization of conserved motifs in HIV-1 Vif required for APOBEC3G and APOBEC3F interaction.J Mol Biol. 2008 Sep 12;381(4):1000-11. doi: 10.1016/j.jmb.2008.06.061. Epub 2008 Jun 28. J Mol Biol. 2008. PMID: 18619467
-
The role of Vif oligomerization and RNA chaperone activity in HIV-1 replication.Virus Res. 2012 Nov;169(2):361-76. doi: 10.1016/j.virusres.2012.06.018. Epub 2012 Jun 21. Virus Res. 2012. PMID: 22728817 Review.
-
Structural disorder in the HIV-1 Vif protein and interaction-dependent gain of structure.Protein Pept Lett. 2010 Aug;17(8):988-98. doi: 10.2174/092986610791498876. Protein Pept Lett. 2010. PMID: 20450485 Review.
Cited by
-
Essential roles of Leu/Ile/Phe-rich domain of JC virus agnoprotein in dimer/oligomer formation, protein stability and splicing of viral transcripts.Virology. 2013 Aug 15;443(1):161-76. doi: 10.1016/j.virol.2013.05.003. Epub 2013 Jun 6. Virology. 2013. PMID: 23747198 Free PMC article.
-
A New Class of Antiretroviral Enabling Innate Immunity by Protecting APOBEC3 from HIV Vif-Dependent Degradation.Trends Mol Med. 2018 May;24(5):507-520. doi: 10.1016/j.molmed.2018.03.004. Epub 2018 Mar 30. Trends Mol Med. 2018. PMID: 29609878 Free PMC article. Review.
-
Codon pairs of the HIV-1 vif gene correlate with CD4+ T cell count.BMC Infect Dis. 2013 Apr 11;13:173. doi: 10.1186/1471-2334-13-173. BMC Infect Dis. 2013. PMID: 23578255 Free PMC article.
-
On the solution conformation and dynamics of the HIV-1 viral infectivity factor.J Mol Biol. 2011 Jul 29;410(5):1008-22. doi: 10.1016/j.jmb.2011.04.053. J Mol Biol. 2011. PMID: 21763503 Free PMC article.
-
Induction of Rod-Shaped Structures by Herpes Simplex Virus Glycoprotein I.J Virol. 2020 Aug 17;94(17):e00231-20. doi: 10.1128/JVI.00231-20. Print 2020 Aug 17. J Virol. 2020. PMID: 32581097 Free PMC article.
References
-
- Malim MH, Emerman M. HIV-1 accessory proteins–ensuring viral survival in a hostile environment. Cell Host Microbe. 2008;3:388–398. - PubMed
-
- Sodroski J, Goh WC, Rosen C, Tartar A, Portetelle D, Burny A, Haseltine W. Replicative and cytopathic potential of HTLV-III/LAV with sor gene deletions. Science. 1986;231:1549–1553. - PubMed
-
- Courcoul M, Patience C, Rey F, Blanc D, Harmache A, Sire J, Vigne R, Spire B. Peripheral blood mononuclear cells produce normal amounts of defective Vif- human immunodeficiency virus type 1 particles which are restricted for the preretrotranscription steps. J. Virol. 1995;69:2068–2074. - PMC - PubMed
