

Structure of a bacterial ribonuclease P holoenzyme in complex with tRNA
- PMID: 21076397
- PMCID: PMC3058908
- DOI: 10.1038/nature09516
Structure of a bacterial ribonuclease P holoenzyme in complex with tRNA
Abstract
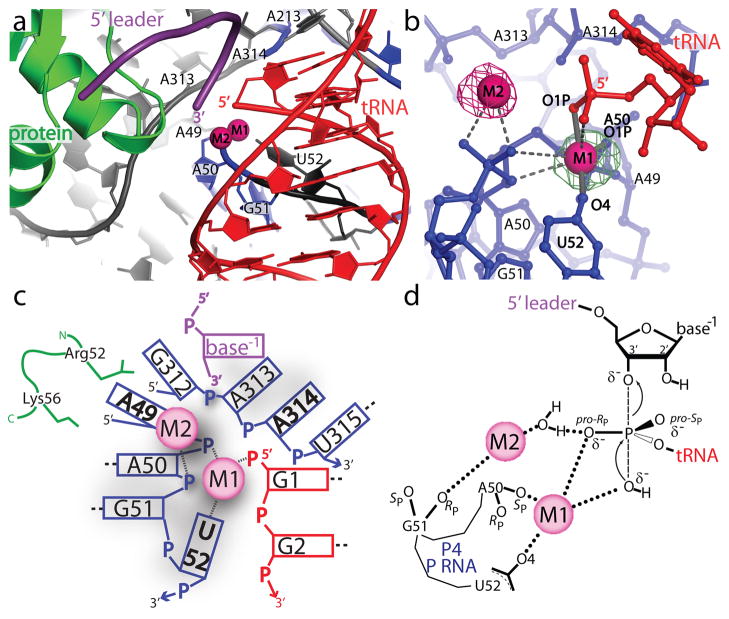
Ribonuclease (RNase) P is the universal ribozyme responsible for 5'-end tRNA processing. We report the crystal structure of the Thermotoga maritima RNase P holoenzyme in complex with tRNA(Phe). The 154 kDa complex consists of a large catalytic RNA (P RNA), a small protein cofactor and a mature tRNA. The structure shows that RNA-RNA recognition occurs through shape complementarity, specific intermolecular contacts and base-pairing interactions. Soaks with a pre-tRNA 5' leader sequence with and without metal help to identify the 5' substrate path and potential catalytic metal ions. The protein binds on top of a universally conserved structural module in P RNA and interacts with the leader, but not with the mature tRNA. The active site is composed of phosphate backbone moieties, a universally conserved uridine nucleobase, and at least two catalytically important metal ions. The active site structure and conserved RNase P-tRNA contacts suggest a universal mechanism of catalysis by RNase P.
Conflict of interest statement
Figures
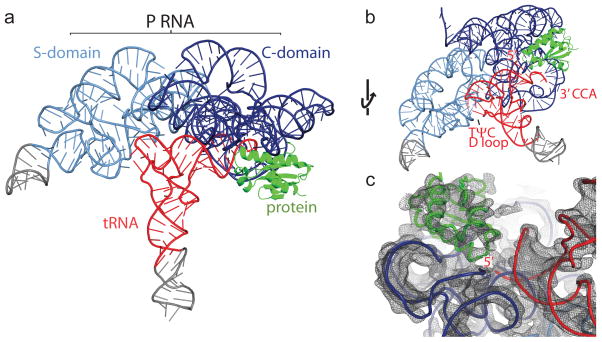
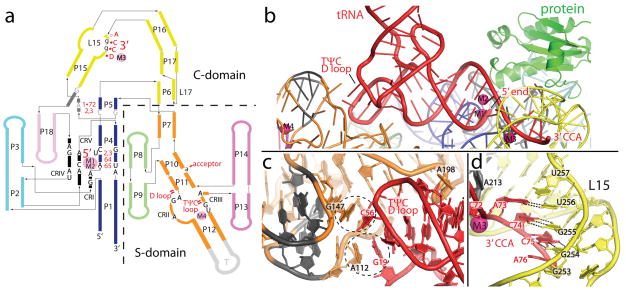
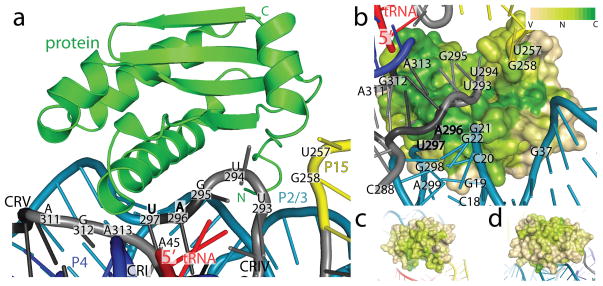
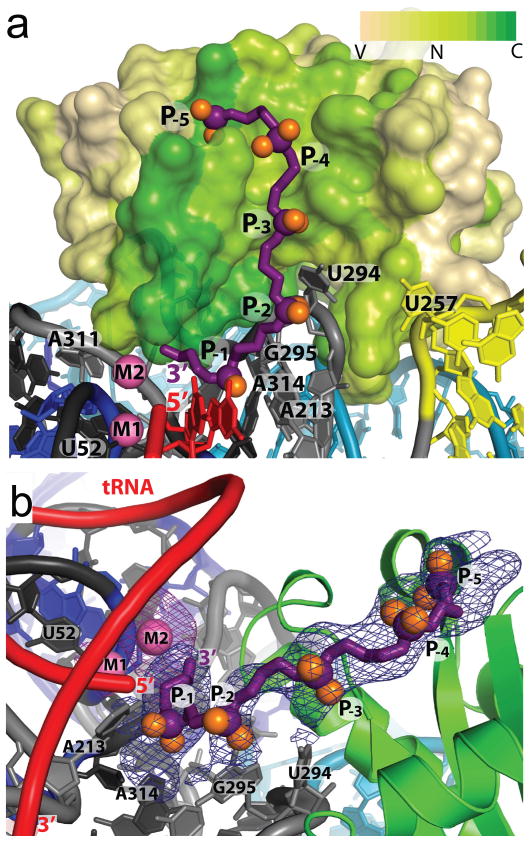
Comment in
-
The RNP bridge between two worlds.Nat Rev Mol Cell Biol. 2011 Mar;12(3):135. doi: 10.1038/nrm3061. Epub 2011 Feb 2. Nat Rev Mol Cell Biol. 2011. PMID: 21285979 No abstract available.
References
-
- Hartmann RK, Gossringer M, Spath B, Fischer S, Marchfelder A. The making of tRNAs and more - RNase P and tRNase Z. Prog Mol Biol Transl Sci. 2009;85:319–368. - PubMed
-
- Kazantsev AV, Pace NR. Bacterial RNase P: a new view of an ancient enzyme. Nat Rev Microbiol. 2006;4:729–740. - PubMed
-
- Liu F, Altman S. Ribonuclease P. Vol. 10. Springer Science+Business Media, LLC; 2010.
-
- Guerrier-Takada C, Gardiner K, Marsh T, Pace N, Altman S. The RNA moiety of ribonuclease P is the catalytic subunit of the enzyme. Cell. 1983;35:849–857. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
