

Translation initiation: variations in the mechanism can be anticipated
- PMID: 21076851
- PMCID: PMC11115079
- DOI: 10.1007/s00018-010-0588-z
Translation initiation: variations in the mechanism can be anticipated
Abstract
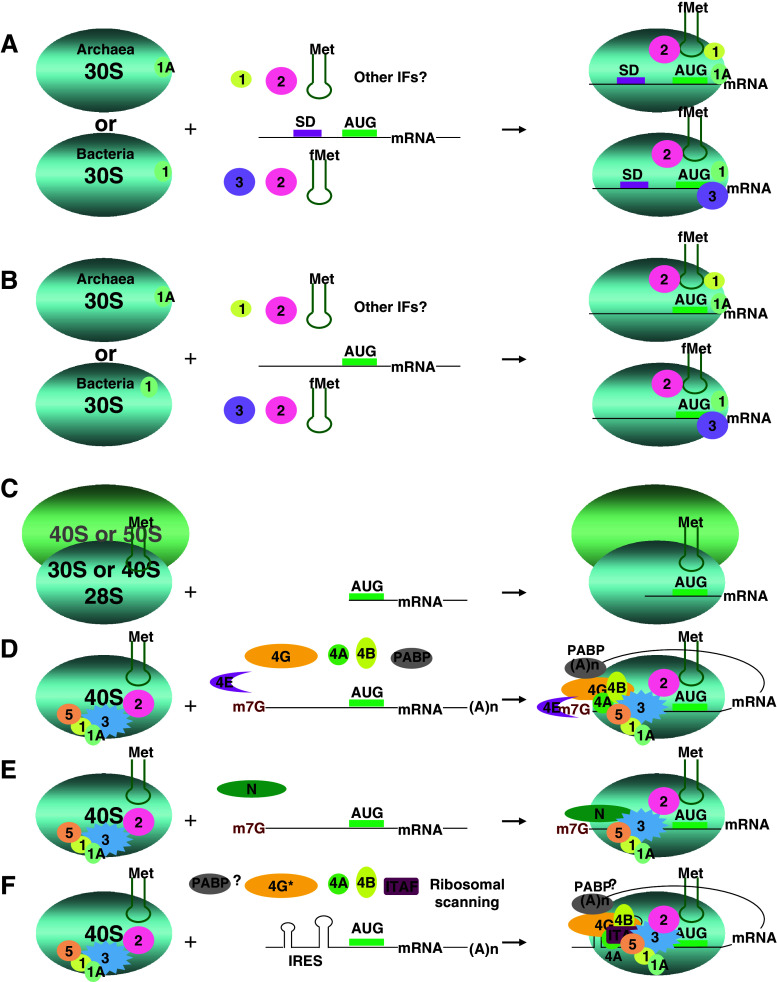
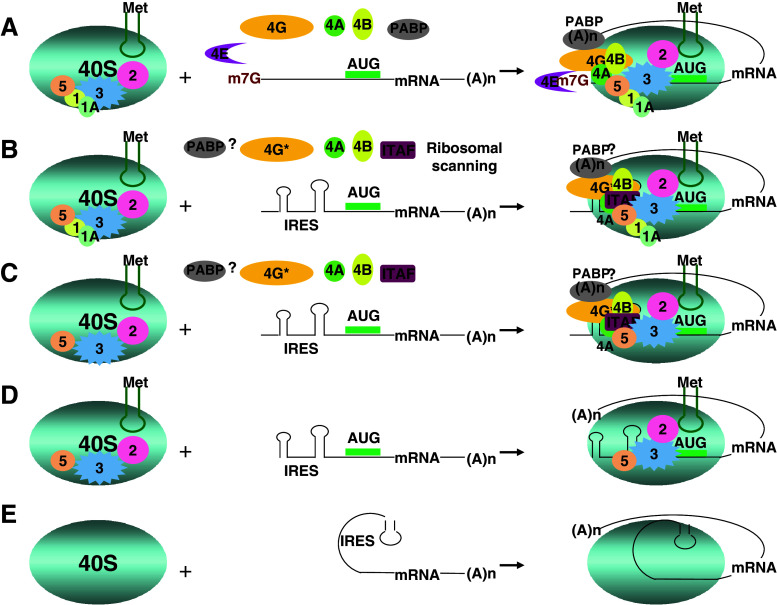
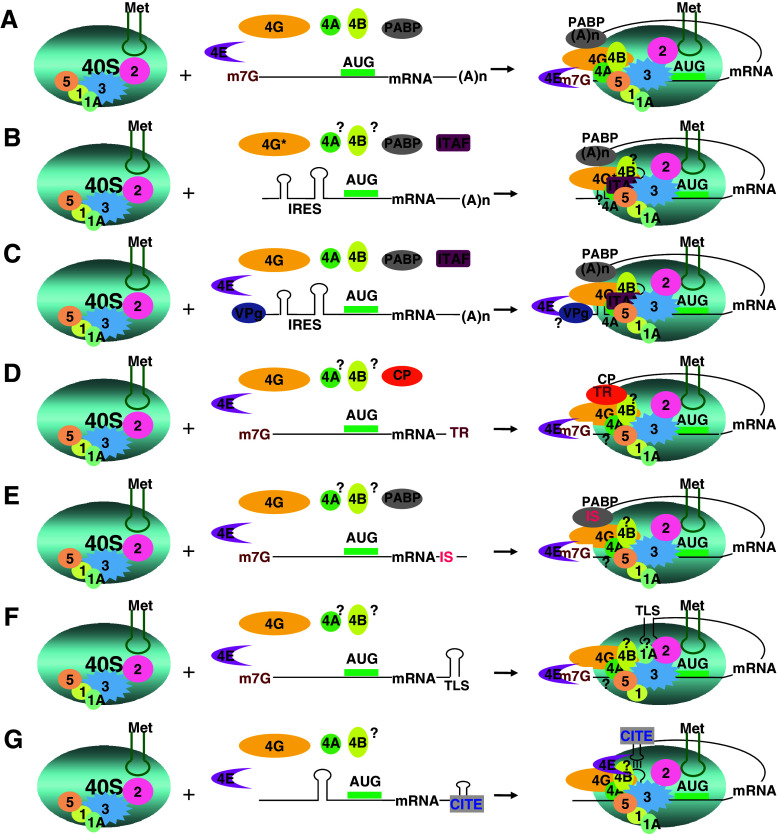
Translation initiation is a critical step in protein synthesis. Previously, two major mechanisms of initiation were considered as essential: prokaryotic, based on SD interaction; and eukaryotic, requiring cap structure and ribosomal scanning. Although discovered decades ago, cap-independent translation has recently been acknowledged as a widely spread mechanism in viruses, which may take place in some cellular mRNA translations. Moreover, it has become evident that translation can be initiated on the leaderless mRNA in all three domains of life. New findings demonstrate that other distinguishable types of initiation exist, including SD-independent in Bacteria and Archaea, and various modifications of 5' end-dependent and internal initiation mechanisms in Eukarya. Since translation initiation has developed through the loss, acquisition, and modification of functional elements, all of which have been elevated by competition with viral translation in a large number of organisms of different complexity, more variation in initiation mechanisms can be anticipated.
Figures
References
-
- Londei P. Translation. In: Cavicchioli R, editor. Archaea: cellular and molecular biology. Washington: ASM Press; 2007. pp. 175–208.
Publication types
MeSH terms
Substances
Grants and funding
- BB/E003729/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/E003192/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/E024181/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/E011888/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/C008219/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
