

Eph/ephrin molecules--a hub for signaling and endocytosis
- PMID: 21078817
- PMCID: PMC2975924
- DOI: 10.1101/gad.1973910
Eph/ephrin molecules--a hub for signaling and endocytosis
Abstract
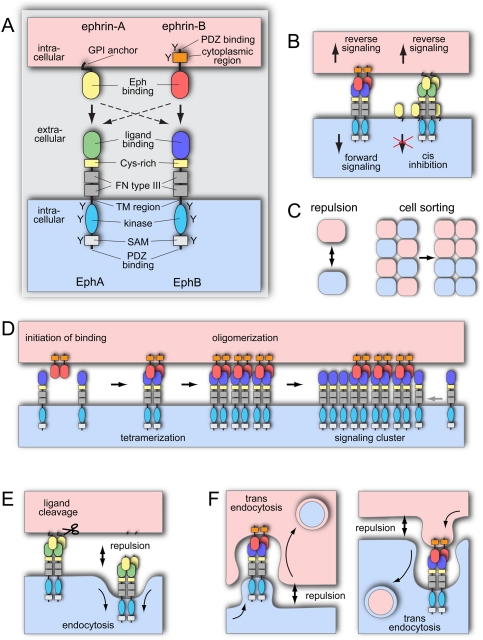
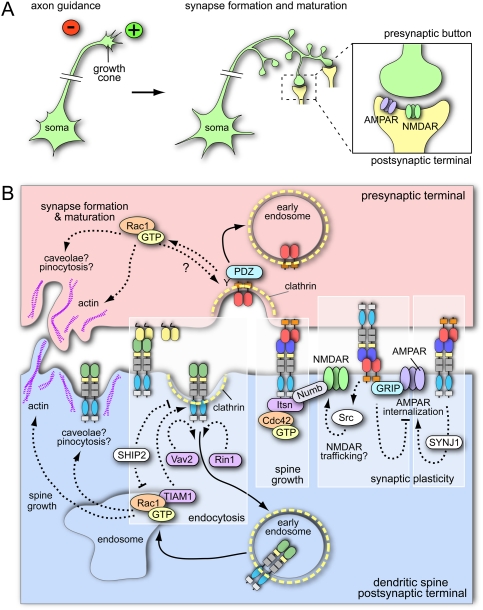
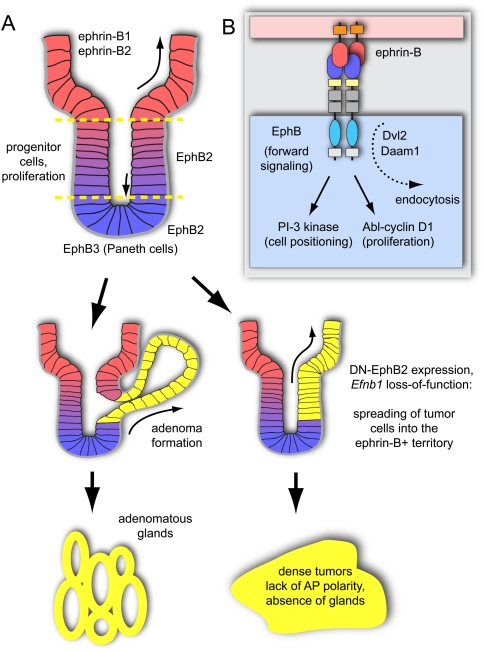
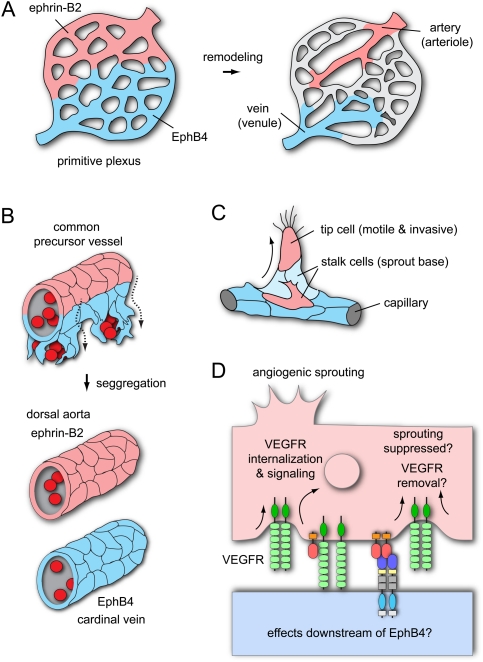
The development, homeostasis, and regeneration of complex organ systems require extensive cell-cell communication to ensure that different cells proliferate, migrate, differentiate, assemble, and function in a coordinated and timely fashion. Eph receptor tyrosine kinases and their ephrin ligands are critical regulators of cell contact-dependent signaling and patterning. Eph/ephrin binding can lead to very diverse biological readouts such as adhesion versus repulsion, or increased versus decreased motility. Accordingly, depending on cell type and context, a limited and conserved set of receptor-ligand interactions is translated into a large variety of downstream signaling processes. Recent evidence indicates that the endocytosis of Eph/ephrin molecules, together with the internalization of various associated tissue-specific effectors, might be one of the key principles responsible for such highly diverse and adaptable biological roles. Here, we summarize recent insights into Eph/ephrin signaling and endocytosis in three biological systems; i.e., the brain, intestine, and vasculature.
Figures
References
-
- Batlle E, Henderson JT, Beghtel H, van den Born MM, Sancho E, Huls G, Meeldijk J, Robertson J, van de Wetering M, Pawson T, et al. 2002. β-Catenin and TCF mediate cell positioning in the intestinal epithelium by controlling the expression of EphB/ephrinB. Cell 111: 251–263 - PubMed
-
- Batlle E, Bacani J, Begthel H, Jonkheer S, Gregorieff A, van de Born M, Malats N, Sancho E, Boon E, Pawson T, et al. 2005. EphB receptor activity suppresses colorectal cancer progression. Nature 435: 1126–1130 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous