

Conflict between noise and plasticity in yeast
- PMID: 21079670
- PMCID: PMC2973811
- DOI: 10.1371/journal.pgen.1001185
Conflict between noise and plasticity in yeast
Abstract
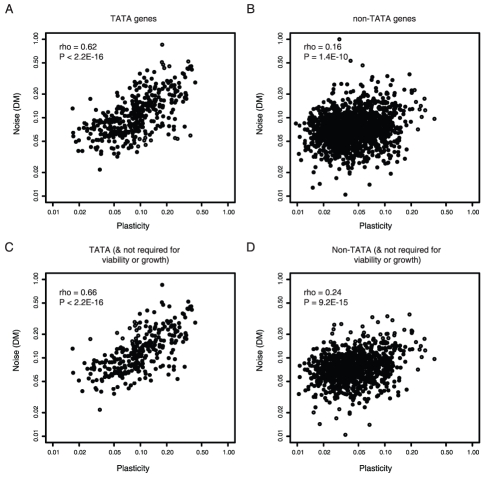
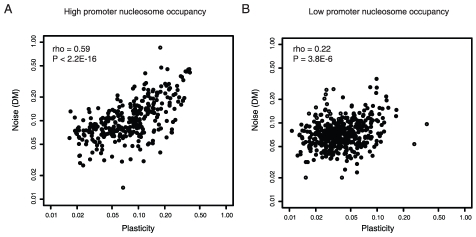
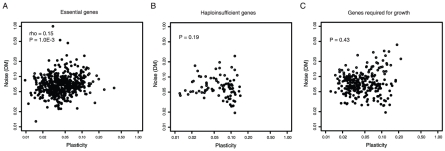
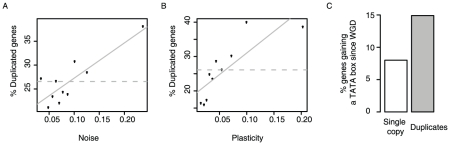
Gene expression responds to changes in conditions but also stochastically among individuals. In budding yeast, both expression responsiveness across conditions ("plasticity") and cell-to-cell variation ("noise") have been quantified for thousands of genes and found to correlate across genes. It has been argued therefore that noise and plasticity may be strongly coupled and mechanistically linked. This is consistent with some theoretical ideas, but a strong coupling between noise and plasticity also has the potential to introduce cost-benefit conflicts during evolution. For example, if high plasticity is beneficial (genes need to respond to the environment), but noise is detrimental (fluctuations are harmful), then strong coupling should be disfavored. Here, evidence is presented that cost-benefit conflicts do occur and that they constrain the evolution of gene expression and promoter usage. In contrast to recent assertions, coupling between noise and plasticity is not a general property, but one associated with particular mechanisms of transcription initiation. Further, promoter architectures associated with coupling are avoided when noise is most likely to be detrimental, and noise and plasticity are largely independent traits for core cellular components. In contrast, when genes are duplicated noise-plasticity coupling increases, consistent with reduced detrimental affects of expression variation. Noise-plasticity coupling is, therefore, an evolvable trait that may constrain the emergence of highly responsive gene expression and be selected against during evolution. Further, the global quantitative data in yeast suggest that one mechanism that relieves the constraints imposed by noise-plasticity coupling is gene duplication, providing an example of how duplication can facilitate escape from adaptive conflicts.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Comment in
-
Noise versus plasticity.Nat Rev Genet. 2011 Jan;12(1):4. doi: 10.1038/nrg2925. Epub 2010 Nov 30. Nat Rev Genet. 2011. PMID: 21116308 No abstract available.
Similar articles
-
Coupling between noise and plasticity in E. coli.G3 (Bethesda). 2013 Dec 9;3(12):2115-20. doi: 10.1534/g3.113.008540. G3 (Bethesda). 2013. PMID: 24122054 Free PMC article.
-
Expression properties exhibit correlated patterns with the fate of duplicated genes, their divergence, and transcriptional plasticity in Saccharomycotina.DNA Res. 2017 Dec 1;24(6):559-570. doi: 10.1093/dnares/dsx025. DNA Res. 2017. PMID: 28633360 Free PMC article.
-
Balancing noise and plasticity in eukaryotic gene expression.BMC Genomics. 2012 Jul 27;13:343. doi: 10.1186/1471-2164-13-343. BMC Genomics. 2012. PMID: 22839658 Free PMC article.
-
Selection for more of the same product as a force to enhance concerted evolution of duplicated genes.Trends Genet. 2006 Dec;22(12):642-4. doi: 10.1016/j.tig.2006.09.014. Epub 2006 Oct 11. Trends Genet. 2006. PMID: 17045359 Review.
-
Cell cycle control of DNA synthesis in budding yeast.Nucleic Acids Res. 1992 May 25;20(10):2403-10. doi: 10.1093/nar/20.10.2403. Nucleic Acids Res. 1992. PMID: 1598198 Free PMC article. Review. No abstract available.
Cited by
-
Phenotype Uniformity in Combined-Stress Environments has a Different Genetic Architecture than in Single-Stress Treatments.Front Plant Sci. 2011 May 4;2:12. doi: 10.3389/fpls.2011.00012. eCollection 2011. Front Plant Sci. 2011. PMID: 22645526 Free PMC article.
-
Broad distribution spectrum from Gaussian to power law appears in stochastic variations in RNA-seq data.Sci Rep. 2018 May 29;8(1):8339. doi: 10.1038/s41598-018-26735-4. Sci Rep. 2018. PMID: 29844539 Free PMC article.
-
Intrinsic adaptive value and early fate of gene duplication revealed by a bottom-up approach.Elife. 2018 Jan 5;7:e29739. doi: 10.7554/eLife.29739. Elife. 2018. PMID: 29303479 Free PMC article.
-
Whole-genome duplication in an algal symbiont bolsters coral heat tolerance.Sci Adv. 2024 Jul 19;10(29):eadn2218. doi: 10.1126/sciadv.adn2218. Epub 2024 Jul 19. Sci Adv. 2024. PMID: 39028812 Free PMC article.
-
DNA structural features of eukaryotic TATA-containing and TATA-less promoters.FEBS Open Bio. 2017 Feb 16;7(3):324-334. doi: 10.1002/2211-5463.12166. eCollection 2017 Mar. FEBS Open Bio. 2017. PMID: 28286728 Free PMC article.
References
-
- Ihmels J, Friedlander G, Bergmann S, Sarig O, Ziv Y, et al. Revealing modular organization in the yeast transcriptional network. Nat Genet. 2002;31:370–377. - PubMed
-
- Newman JR, Ghaemmaghami S, Ihmels J, Breslow DK, Noble M, et al. Single-cell proteomic analysis of S. cerevisiae reveals the architecture of biological noise. Nature. 2006;441:840–846. - PubMed
-
- Choi JK, Kim YJ. Intrinsic variability of gene expression encoded in nucleosome positioning sequences. Nat Genet. 2009;41:498–503. - PubMed
-
- Landry CR, Lemos B, Rifkin SA, Dickinson WJ, Hartl DL. Genetic properties influencing the evolvability of gene expression. Science. 2007;317:118–121. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
