

Translation elongation factor 1A facilitates the assembly of the tombusvirus replicase and stimulates minus-strand synthesis
- PMID: 21079685
- PMCID: PMC2973826
- DOI: 10.1371/journal.ppat.1001175
Translation elongation factor 1A facilitates the assembly of the tombusvirus replicase and stimulates minus-strand synthesis
Abstract
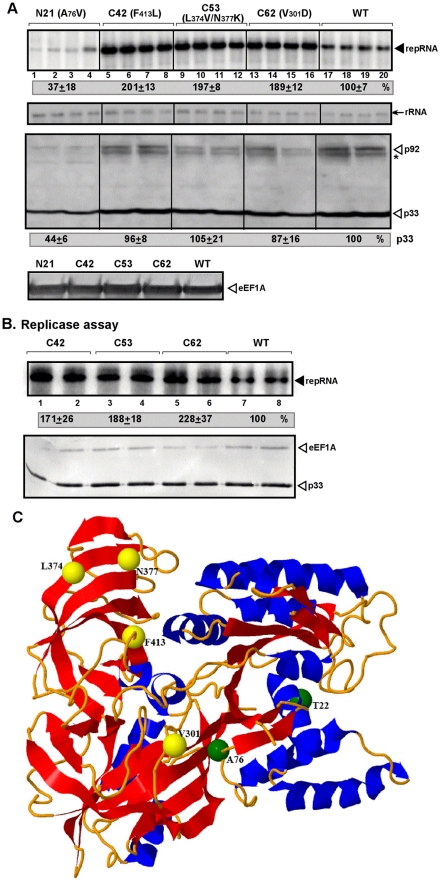
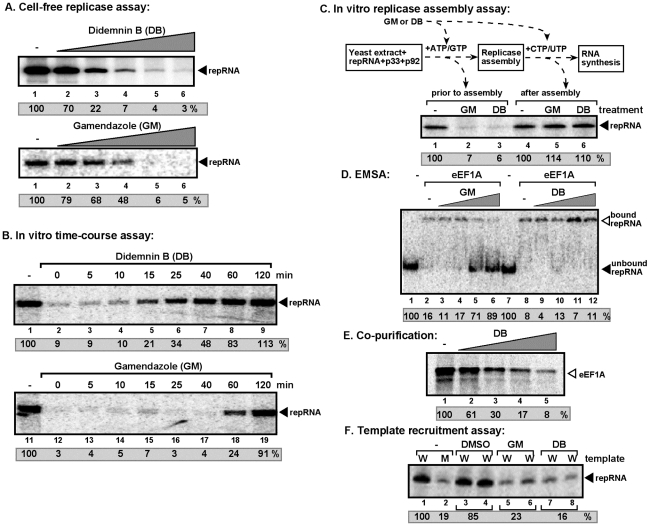
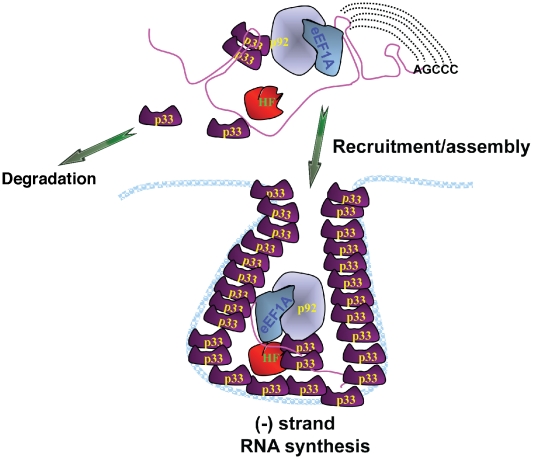
Replication of plus-strand RNA viruses depends on host factors that are recruited into viral replicase complexes. Previous studies showed that eukaryotic translation elongation factor (eEF1A) is one of the resident host proteins in the highly purified tombusvirus replicase complex. Using a random library of eEF1A mutants, we identified one mutant that decreased and three mutants that increased Tomato bushy stunt virus (TBSV) replication in a yeast model host. Additional in vitro assays with whole cell extracts prepared from yeast strains expressing the eEF1A mutants demonstrated several functions for eEF1A in TBSV replication: facilitating the recruitment of the viral RNA template into the replicase complex; the assembly of the viral replicase complex; and enhancement of the minus-strand synthesis by promoting the initiation step. These roles for eEF1A are separate from its canonical role in host and viral protein translation, emphasizing critical functions for this abundant cellular protein during TBSV replication.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
