

Perfringolysin O association with ordered lipid domains: implications for transmembrane protein raft affinity
- PMID: 21081073
- PMCID: PMC2980714
- DOI: 10.1016/j.bpj.2010.09.028
Perfringolysin O association with ordered lipid domains: implications for transmembrane protein raft affinity
Abstract
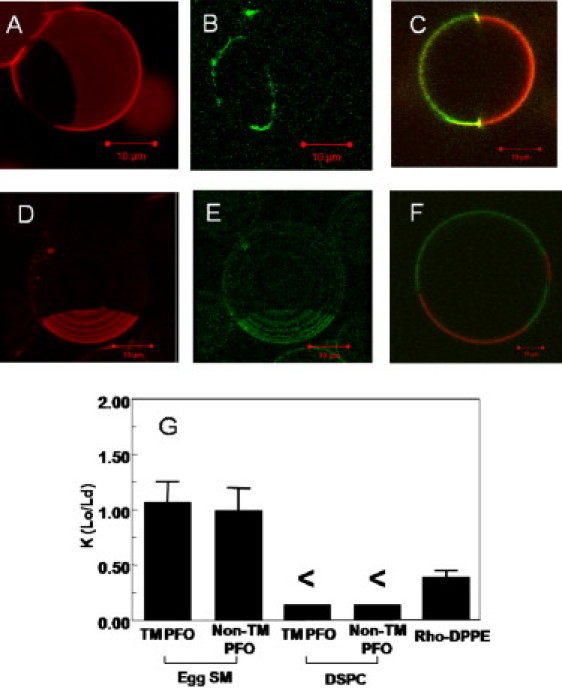
Upon interaction with cholesterol, perfringolysin O (PFO) inserts into membranes and forms a rigid transmembrane (TM) β-barrel. PFO is believed to interact with liquid ordered lipid domains (lipid rafts). Because the origin of TM protein affinity for rafts is poorly understood, we investigated PFO raft affinity in vesicles having coexisting ordered and disordered lipid domains. Fluorescence resonance energy transfer (FRET) from PFO Trp to domain-localized acceptors indicated that PFO generally has a raft affinity between that of LW peptide (low raft affinity) and cholera toxin B (high raft affinity) in vesicles containing ordered domains rich in brain sphingomyelin or distearoylphosphatidylcholine. FRET also showed that ceramide, which increases exposure of cholesterol to water and thus displaces it from rafts, does not displace PFO from ordered domains. This can be explained by shielding of PFO-bound cholesterol from water. Finally, FRET showed that PFO affinity for ordered domains was higher in its non-TM (prepore) form than in its TM form, demonstrating that the TM portion of PFO interacts unfavorably with rafts. Microscopy studies in giant unilamellar vesicles confirmed that PFO exhibits intermediate raft affinity, and showed that TM PFO (but not non-TM PFO) concentrated at the edges of liquid ordered domains. These studies suggest that a combination of binding to raft-associating molecules and having a rigid TM structure that is unable to pack well in a highly ordered lipid environment can control TM protein domain localization. To accommodate these constraints, raft-associated TM proteins in cells may tend to locate within liquid disordered shells encapsulated within ordered domains.
Copyright © 2010 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures


References
-
- Tweten R.K., Parker M.W., Johnson A.E. The cholesterol-dependent cytolysins. Curr. Top. Microbiol. Immunol. 2001;257:15–33. - PubMed
-
- London E. How principles of domain formation in model membranes may explain ambiguities concerning lipid raft formation in cells. Biochim. Biophys. Acta. 2005;1746:203–220. - PubMed
-
- Ohno-Iwashita Y., Shimada Y., Iwashita S. Perfringolysin O, a cholesterol-binding cytolysin, as a probe for lipid rafts. Anaerobe. 2004;10:125–134. - PubMed
-
- Shimada Y., Maruya M., Ohno-Iwashita Y. The C-terminal domain of perfringolysin O is an essential cholesterol-binding unit targeting to cholesterol-rich microdomains. Eur. J. Biochem. 2002;269:6195–6203. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
