

Many putative endocrine disruptors inhibit prostaglandin synthesis
- PMID: 21081300
- PMCID: PMC3080937
- DOI: 10.1289/ehp.1002635
Many putative endocrine disruptors inhibit prostaglandin synthesis
Abstract
Background: Prostaglandins (PGs) play key roles in development and maintenance of homeostasis of the adult body. Despite these important roles, it remains unclear whether the PG pathway is a target for endocrine disruption. However, several known endocrine-disrupting compounds (EDCs) share a high degree of structural similarity with mild analgesics.
Objectives and methods: Using cell-based transfection and transduction experiments, mass spectrometry, and organotypic assays together with molecular modeling, we investigated whether inhibition of the PG pathway by known EDCs could be a novel point of endocrine disruption.
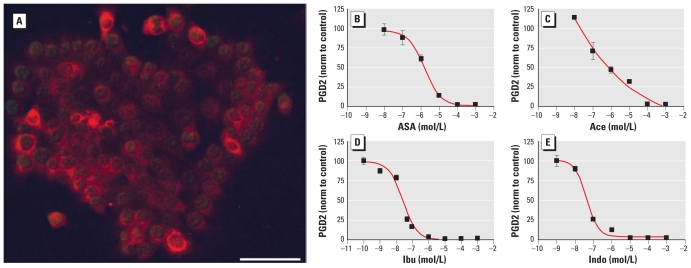
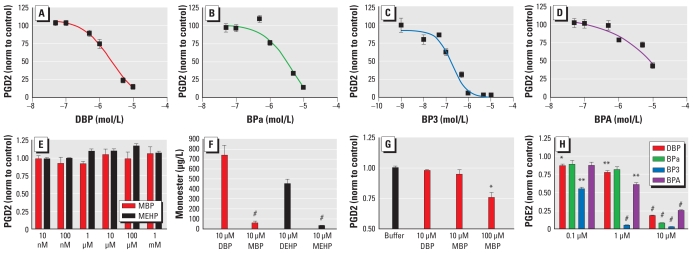
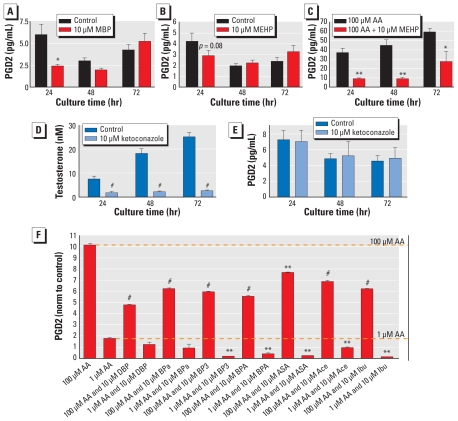
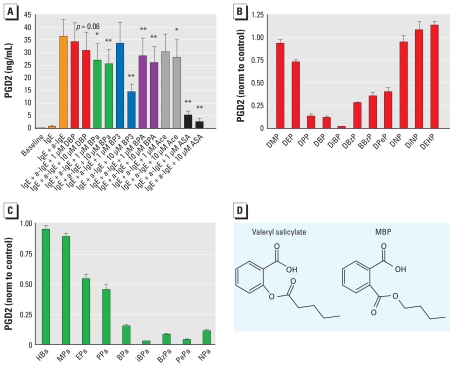
Results: We found that many known EDCs inhibit the PG pathway in a mouse Sertoli cell line and in human primary mast cells. The EDCs also reduced PG synthesis in ex vivo rat testis, and this reduction was correlated with a reduced testosterone production. The inhibition of PG synthesis occurred without involvement of canonical PG receptors or the peroxisome proliferator-activated receptors (PPARs), which have previously been described as targets of EDCs. Instead, our results suggest that the compounds may bind directly into the active site of the cyclooxygenase (COX) enzymes, thereby obstructing the conversion of arachidonic acid to PG precursors without interfering with the expression of the COX enzymes. A common feature of the PG inhibitory EDCs is the presence of aromatic groups that may stabilize binding in the hydrophobic active site of the COX enzymes.
Conclusion: Our findings suggest a hitherto unknown mode of action by EDCs through inhibition of the PG pathway and suggest new avenues to investigate effects of EDCs on reproductive and immunological disorders that have become increasingly common in recent decades.
Figures
References
-
- Adams IR, McLaren A. Sexually dimorphic development of mouse primordial germ cells: switching from oogenesis to spermatogenesis. Development. 2002;129:1155–1164. - PubMed
-
- Amateau SK, McCarthy MM. Induction of PGE2 by estradiol mediates developmental masculinization of sex behavior. Nat Neurosci. 2004;7:643–650. - PubMed
-
- Beasley R, Clayton T, Crane J, von Mutius E, Lai CK, Montefort S, et al. Association between paracetamol use in infancy and childhood, and risk of asthma, rhinoconjunctivitis, and eczema in children aged 6–7 years: analysis from phase three of the ISAAC programme. Lancet. 2008;372:1039–1048. - PubMed
-
- Boisen KA, Kaleva M, Main KM, Virtanen HE, Haavisto AM, Schmidt IM, et al. Difference in prevalence of congenital cryptorchidism in infants between two Nordic countries. Lancet. 2004;363:1264–1269. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
