

Revisiting HIV-1 uncoating
- PMID: 21083892
- PMCID: PMC2998454
- DOI: 10.1186/1742-4690-7-96
Revisiting HIV-1 uncoating
Abstract
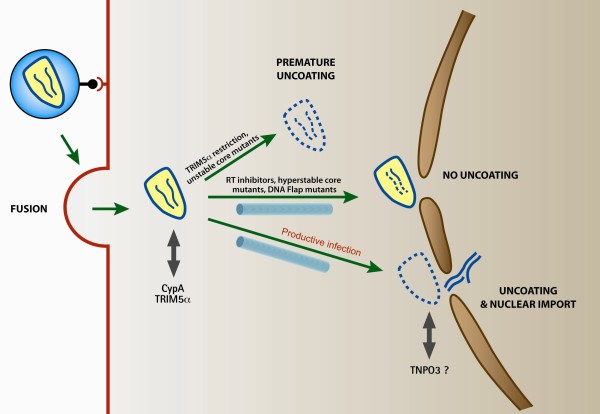
HIV uncoating is defined as the loss of viral capsid that occurs within the cytoplasm of infected cells before entry of the viral genome into the nucleus. It is an obligatory step of HIV-1 early infection and accompanies the transition between reverse transcription complexes (RTCs), in which reverse transcription occurs, and pre-integration complexes (PICs), which are competent to integrate into the host genome. The study of the nature and timing of HIV-1 uncoating has been paved with difficulties, particularly as a result of the vulnerability of the capsid assembly to experimental manipulation. Nevertheless, recent studies of capsid structure, retroviral restriction and mechanisms of nuclear import, as well as the recent expansion of technical advances in genome-wide studies and cell imagery approaches, have substantially changed our understanding of HIV uncoating. Although early work suggested that uncoating occurs immediately following viral entry in the cell, thus attributing a trivial role for the capsid in infected cells, recent data suggest that uncoating occurs several hours later and that capsid has an all-important role in the cell that it infects: for transport towards the nucleus, reverse transcription and nuclear import. Knowing that uncoating occurs at a later stage suggests that the viral capsid interacts extensively with the cytoskeleton and other cytoplasmic components during its transport to the nucleus, which leads to a considerable reassessment of our efforts to identify potential therapeutic targets for HIV therapy. This review discusses our current understanding of HIV uncoating, the functional interplay between infectivity and timely uncoating, as well as exposing the appropriate methods to study uncoating and addressing the many questions that remain unanswered.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
