

Structural and biophysical analysis of BST-2/tetherin ectodomains reveals an evolutionary conserved design to inhibit virus release
- PMID: 21084286
- PMCID: PMC3024793
- DOI: 10.1074/jbc.M110.190538
Structural and biophysical analysis of BST-2/tetherin ectodomains reveals an evolutionary conserved design to inhibit virus release
Abstract
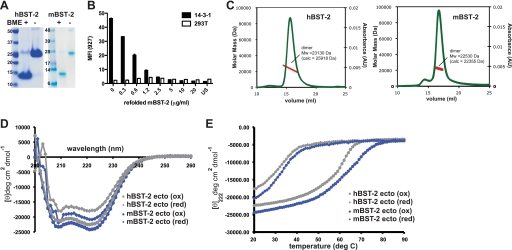
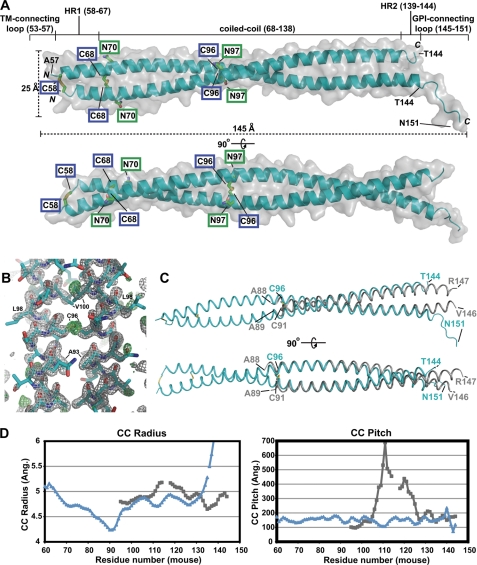
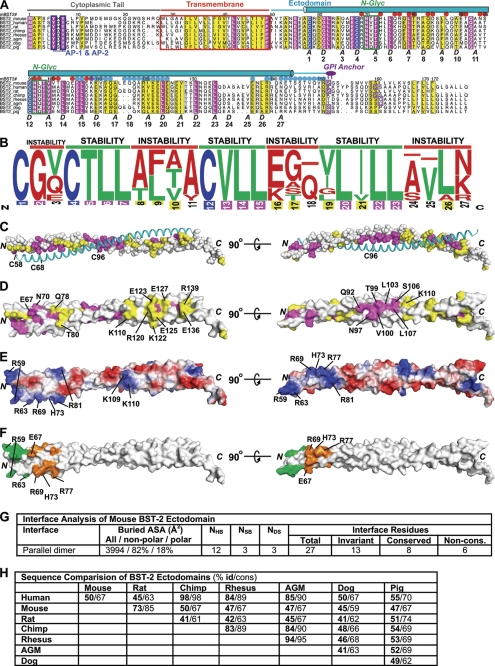
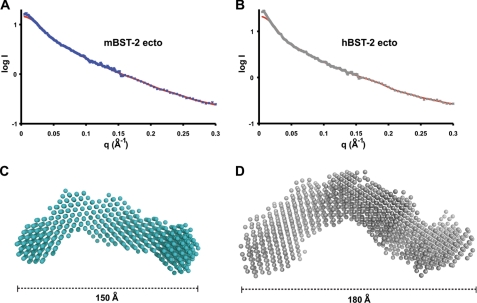
BST-2/tetherin is a host antiviral molecule that functions to potently inhibit the release of enveloped viruses from infected cells. In return, viruses have evolved antagonists to this activity. BST-2 traps budding virions by using two separate membrane-anchoring regions that simultaneously incorporate into the host and viral membranes. Here, we detailed the structural and biophysical properties of the full-length BST-2 ectodomain, which spans the two membrane anchors. The 1.6-Å crystal structure of the complete mouse BST-2 ectodomain reveals an ∼145-Å parallel dimer in an extended α-helix conformation that predominantly forms a coiled coil bridged by three intermolecular disulfides that are required for stability. Sequence analysis in the context of the structure revealed an evolutionarily conserved design that destabilizes the coiled coil, resulting in a labile superstructure, as evidenced by solution x-ray scattering displaying bent conformations spanning 150 and 180 Å for the mouse and human BST-2 ectodomains, respectively. Additionally, crystal packing analysis revealed possible curvature-sensing tetrameric structures that may aid in proper placement of BST-2 during the genesis of viral progeny. Overall, this extended coiled-coil structure with inherent plasticity is undoubtedly necessary to accommodate the dynamics of viral budding while ensuring separation of the anchors.
Figures
References
-
- Sauter D., Specht A., Kirchhoff F. (2010) Cell 141, 392–398 - PubMed
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
