

Cell-based analysis of structure-function activity of threonine aspartase 1
- PMID: 21084304
- PMCID: PMC3024795
- DOI: 10.1074/jbc.M110.161646
Cell-based analysis of structure-function activity of threonine aspartase 1
Abstract
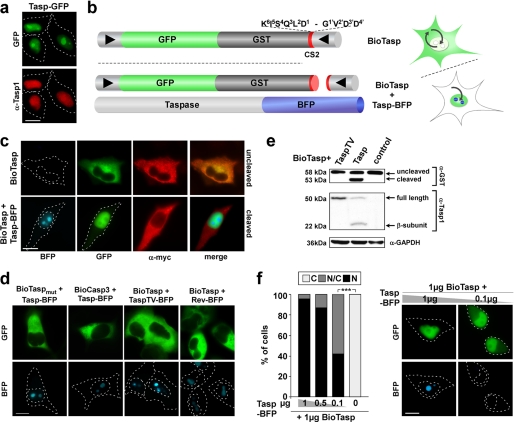
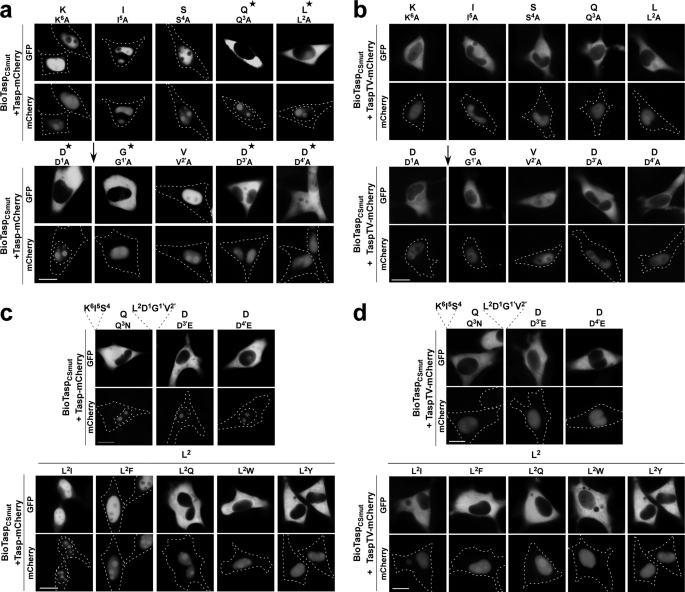
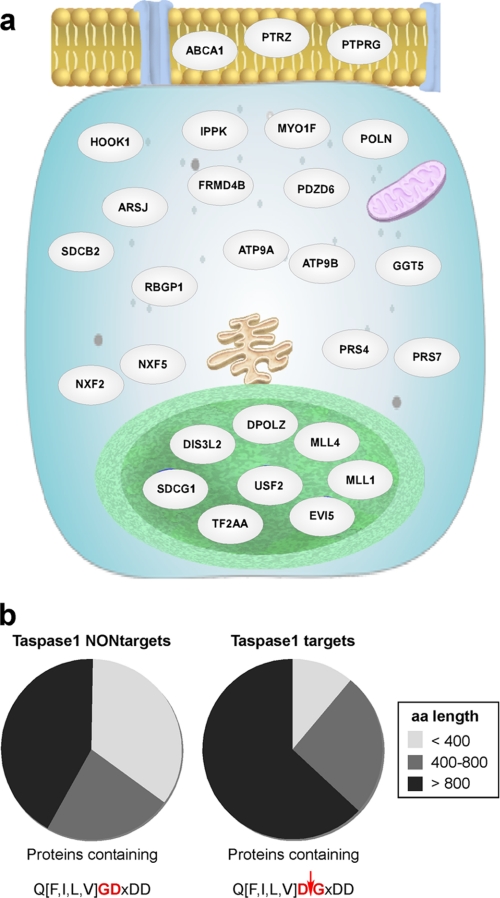
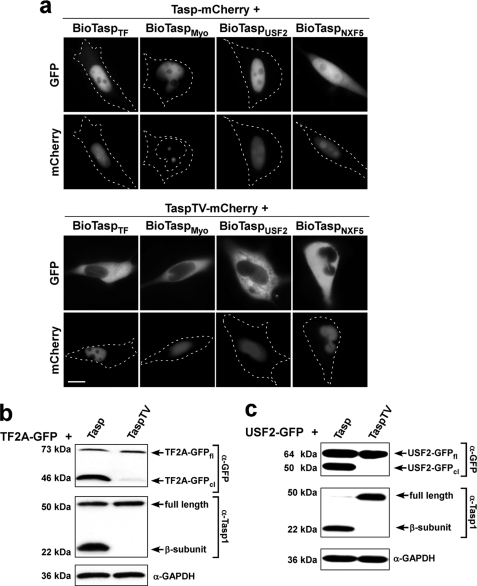
Taspase1 is a threonine protease responsible for cleaving intracellular substrates. As such, (de)regulated Taspase1 function is expected not only to be vital for ordered development but may also be relevant for disease. However, the full repertoires of Taspase1 targets as well as the exact biochemical requirements for its efficient and substrate-specific cleavage are not yet resolved. Also, no cellular assays for this protease are currently available, hampering the exploitation of the (patho)biological relevance of Taspase1. Here, we developed highly efficient cell-based translocation biosensor assays to probe Taspase1 trans-cleavage in vivo. These modular sensors harbor variations of Taspase1 cleavage sites and localize to the cytoplasm. Expression of Taspase1 but not of inactive Taspase1 mutants or of unrelated proteases triggers proteolytic cleavage and nuclear accumulation of the biosensors. Employing our assay combined with scanning mutagenesis, we identified the sequence and spatial requirements for efficient Taspase1 processing in liquid and solid tumor cell lines. Collectively, our results defined an improved Taspase1 consensus recognition sequence, Q(3)(F/I/L/V)(2)D(1)↓G(1)'X(2)'D(3)'D(4)', allowing the first genome-wide bioinformatic identification of the human Taspase1 degradome. Among the 27 most likely Taspase1 targets are cytoplasmic but also nuclear proteins, such as the upstream stimulatory factor 2 (USF2) or the nuclear RNA export factors 2/5 (NXF2/5). Cleavage site recognition and proteolytic processing of selected targets were verified in the context of the biosensor and for the full-length proteins. We provide novel mechanistic insights into the function and bona fide targets of Taspase1 allowing for a focused investigation of the (patho)biological relevance of this type 2 asparaginase.
Figures
Similar articles
-
Taspase1: a 'misunderstood' protease with translational cancer relevance.Oncogene. 2016 Jun 30;35(26):3351-64. doi: 10.1038/onc.2015.436. Epub 2015 Dec 14. Oncogene. 2016. PMID: 26657154 Review.
-
Bioassays to monitor Taspase1 function for the identification of pharmacogenetic inhibitors.PLoS One. 2011;6(5):e18253. doi: 10.1371/journal.pone.0018253. Epub 2011 May 25. PLoS One. 2011. PMID: 21647428 Free PMC article.
-
Fly versus man: evolutionary impairment of nucleolar targeting affects the degradome of Drosophila's Taspase1.FASEB J. 2015 May;29(5):1973-85. doi: 10.1096/fj.14-262451. Epub 2015 Jan 29. FASEB J. 2015. PMID: 25634959
-
The importin-alpha/nucleophosmin switch controls taspase1 protease function.Traffic. 2011 Jun;12(6):703-14. doi: 10.1111/j.1600-0854.2011.01191.x. Epub 2011 Apr 13. Traffic. 2011. PMID: 21418451
-
Cleaving for growth: threonine aspartase 1--a protease relevant for development and disease.FASEB J. 2016 Mar;30(3):1012-22. doi: 10.1096/fj.15-270611. Epub 2015 Nov 17. FASEB J. 2016. PMID: 26578689 Review.
Cited by
-
The Vitamin D Receptor-BIM Axis Overcomes Cisplatin Resistance in Head and Neck Cancer.Cancers (Basel). 2022 Oct 19;14(20):5131. doi: 10.3390/cancers14205131. Cancers (Basel). 2022. PMID: 36291915 Free PMC article.
-
Overexpression of the catalytically impaired Taspase1 T234V or Taspase1 D233A variants does not have a dominant negative effect in T(4;11) leukemia cells.PLoS One. 2012;7(5):e34142. doi: 10.1371/journal.pone.0034142. Epub 2012 May 3. PLoS One. 2012. PMID: 22570686 Free PMC article.
-
Disease-relevant signalling-pathways in head and neck cancer: Taspase1's proteolytic activity fine-tunes TFIIA function.Sci Rep. 2017 Nov 2;7(1):14937. doi: 10.1038/s41598-017-14814-x. Sci Rep. 2017. PMID: 29097782 Free PMC article.
-
The protein corona protects against size- and dose-dependent toxicity of amorphous silica nanoparticles.Beilstein J Nanotechnol. 2014 Aug 27;5:1380-92. doi: 10.3762/bjnano.5.151. eCollection 2014. Beilstein J Nanotechnol. 2014. PMID: 25247121 Free PMC article.
-
Taspase1: a 'misunderstood' protease with translational cancer relevance.Oncogene. 2016 Jun 30;35(26):3351-64. doi: 10.1038/onc.2015.436. Epub 2015 Dec 14. Oncogene. 2016. PMID: 26657154 Review.
References
-
- Overall C. M., Dean R. A. (2006) Cancer Metastasis Rev. 25, 69–75 - PubMed
-
- Knauer S. K., Moodt S., Berg T., Liebel U., Pepperkok R., Stauber R. H. (2005) Traffic 6, 594–606 - PubMed
-
- Turk B. (2006) Nat. Rev. Drug Discov. 5, 785–799 - PubMed
-
- Hsieh J. J., Cheng E. H., Korsmeyer S. J. (2003) Cell 115, 293–303 - PubMed
-
- Khan J. A., Dunn B. M., Tong L. (2005) Structure 13, 1443–1452 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
