

The arginine clusters of the carboxy-terminal domain of the core protein of hepatitis B virus make pleiotropic contributions to genome replication
- PMID: 21084467
- PMCID: PMC3020508
- DOI: 10.1128/JVI.01957-10
The arginine clusters of the carboxy-terminal domain of the core protein of hepatitis B virus make pleiotropic contributions to genome replication
Abstract
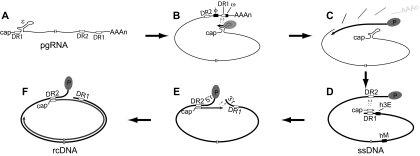
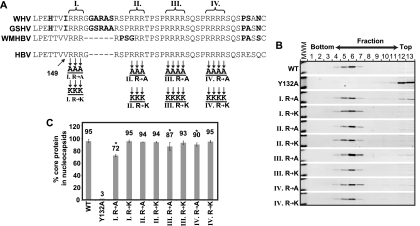
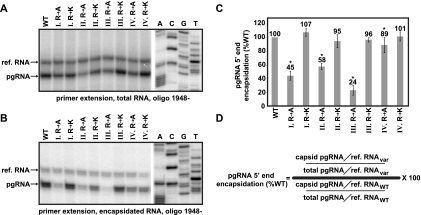
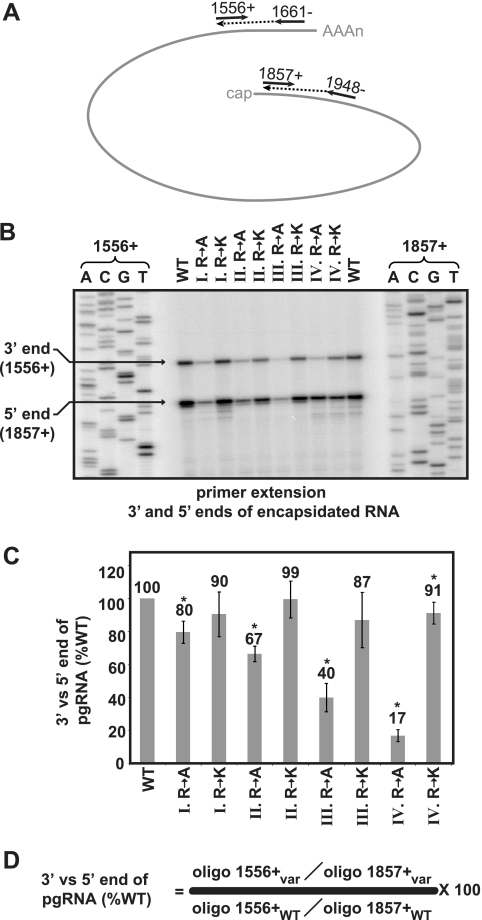
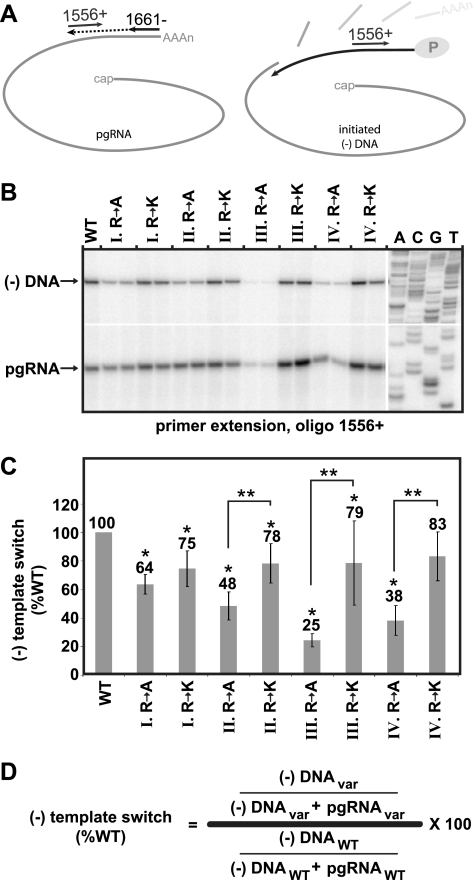
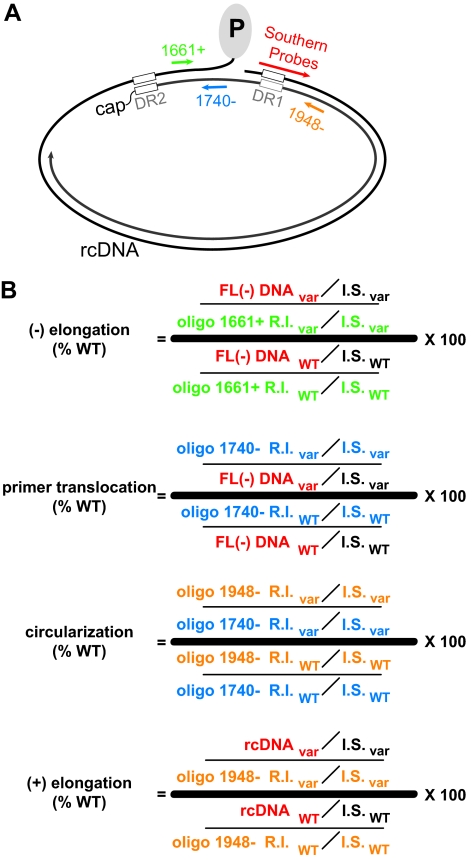
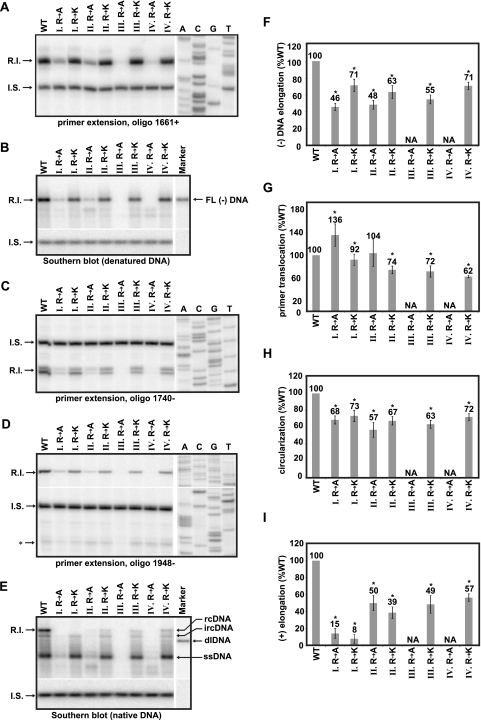
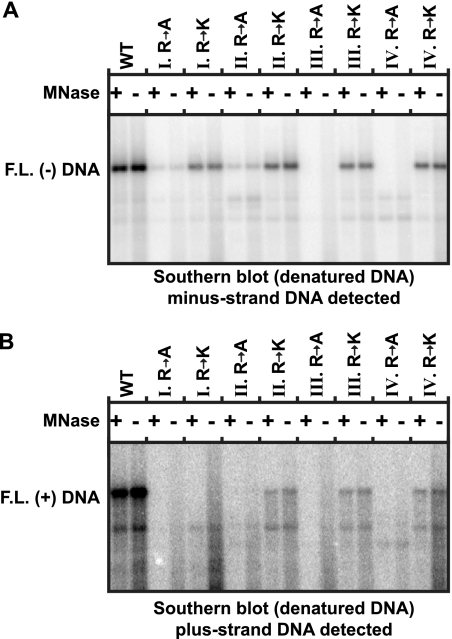
The carboxy-terminal domain (CTD) of the core protein of hepatitis B virus is not necessary for capsid assembly. However, the CTD does contribute to encapsidation of pregenomic RNA (pgRNA). The contribution of the CTD to DNA synthesis is less clear. This is the case because some mutations within the CTD increase the proportion of spliced RNA to pgRNA that are encapsidated and reverse transcribed. The CTD contains four clusters of consecutive arginine residues. The contributions of the individual arginine clusters to genome replication are unknown. We analyzed core protein variants in which the individual arginine clusters were substituted with either alanine or lysine residues. We developed assays to analyze these variants at specific steps throughout genome replication. We used a replication template that was not spliced in order to study the replication of only pgRNA. We found that alanine substitutions caused defects at both early and late steps in genome replication. Lysine substitutions also caused defects, but primarily during later steps. These findings demonstrate that the CTD contributes to DNA synthesis pleiotropically and that preserving the charge within the CTD is not sufficient to preserve function.
Figures
References
-
- Auxilien, S., G. Keith, S. F. Le Grice, and J. L. Darlix. 1999. Role of posttranscriptional modifications of primer tRNALys,3 in the fidelity and efficacy of plus strand DNA transfer during HIV-1 reverse transcription. J. Biol. Chem. 274:4412-4420. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
