

Mapping the landscape of host-pathogen coevolution: HLA class I binding and its relationship with evolutionary conservation in human and viral proteins
- PMID: 21084470
- PMCID: PMC3020499
- DOI: 10.1128/JVI.01966-10
Mapping the landscape of host-pathogen coevolution: HLA class I binding and its relationship with evolutionary conservation in human and viral proteins
Abstract
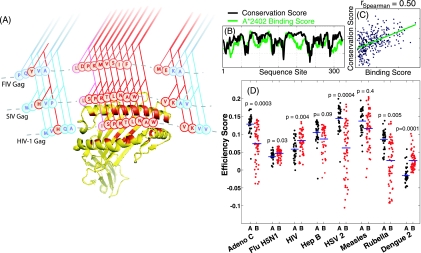
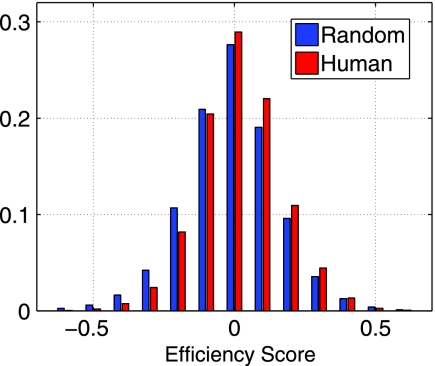
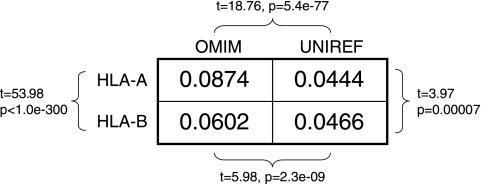
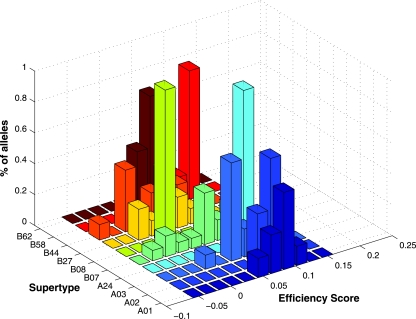
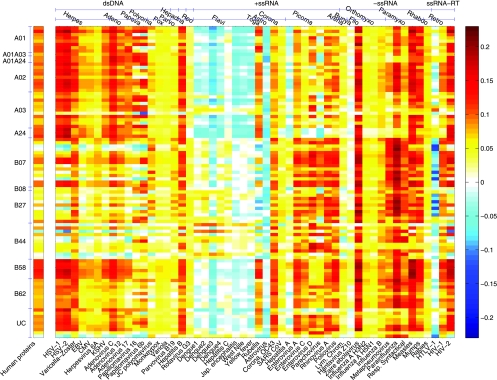
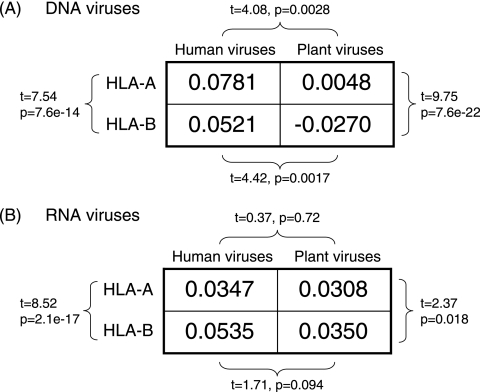
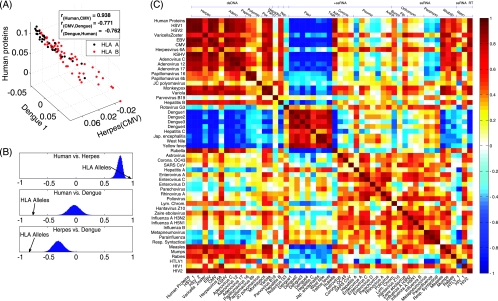
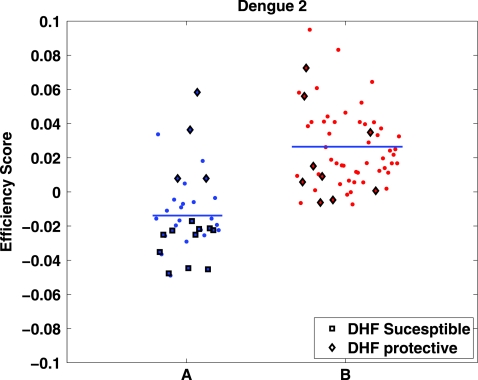
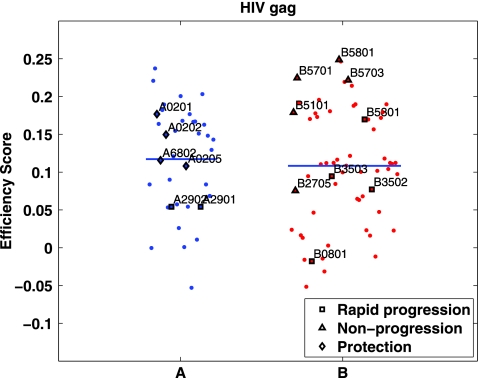
The high diversity of HLA binding preferences has been driven by the sequence diversity of short segments of relevant pathogenic proteins presented by HLA molecules to the immune system. To identify possible commonalities in HLA binding preferences, we quantify these using a novel measure termed "targeting efficiency," which captures the correlation between HLA-peptide binding affinities and the conservation of the targeted proteomic regions. Analysis of targeting efficiencies for 95 HLA class I alleles over thousands of human proteins and 52 human viruses indicates that HLA molecules preferentially target conserved regions in these proteomes, although the arboviral Flaviviridae are a notable exception where nonconserved regions are preferentially targeted by most alleles. HLA-A alleles and several HLA-B alleles that have maintained close sequence identity with chimpanzee homologues target conserved human proteins and DNA viruses such as Herpesviridae and Adenoviridae most efficiently, while all HLA-B alleles studied efficiently target RNA viruses. These patterns of host and pathogen specialization are both consistent with coevolutionary selection and functionally relevant in specific cases; for example, preferential HLA targeting of conserved proteomic regions is associated with improved outcomes in HIV infection and with protection against dengue hemorrhagic fever. Efficiency analysis provides a novel perspective on the coevolutionary relationship between HLA class I molecular diversity, self-derived peptides that shape T-cell immunity through ontogeny, and the broad range of viruses that subsequently engage with the adaptive immune response.
Figures
References
-
- Berezin, C., et al. 2004. ConSeq: the identification of functionally and structurally important residues in protein sequences. Bioinformatics 20:1322-1324. - PubMed
-
- Bhasin, M., and G. P. Raghava. 2007. A hybrid approach for predicting promiscuous MHC class I restricted T cell epitopes. J. Biosci. 32:31-42. - PubMed
-
- Bhattacharya, T., et al. 2007. Founder effects in the assessment of HIV polymorphisms and HLA allele associations. Science 315:1583-1586. - PubMed
-
- Borghans, J. A. M., J. B. Beltman, and R. J. De Boer. 2004. MHC polymorphism under host-pathogen coevolution. Immunogenetics 55:732-739. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
