

Metzincin proteases and their inhibitors: foes or friends in nervous system physiology?
- PMID: 21084591
- PMCID: PMC3072038
- DOI: 10.1523/JNEUROSCI.3467-10.2010
Metzincin proteases and their inhibitors: foes or friends in nervous system physiology?
Abstract
Members of the metzincin family of metalloproteinases have long been considered merely degradative enzymes for extracellular matrix molecules. Recently, however, there has been growing appreciation for these proteinases and their endogenous inhibitors, tissue inhibitors of metalloproteinases (TIMPs), as fine modulators of nervous system physiology and pathology. Present all along the phylogenetic tree, in all neural cell types, from the nucleus to the synapse and in the extracellular space, metalloproteinases exhibit a complex spatiotemporal profile of expression in the nervous parenchyma and at the neurovascular interface. The irreversibility of their proteolytic activity on numerous biofactors (e.g., growth factors, cytokines, receptors, DNA repair enzymes, matrix proteins) is ideally suited to sustain structural changes that are involved in physiological or postlesion remodeling of neural networks, learning consolidation or impairment, neurodegenerative and neuroinflammatory processes, or progression of malignant gliomas. The present review provides a state of the art overview of the involvement of the metzincin/TIMP system in these processes and the prospects of new therapeutic strategies based on the control of metalloproteinase activity.
Figures


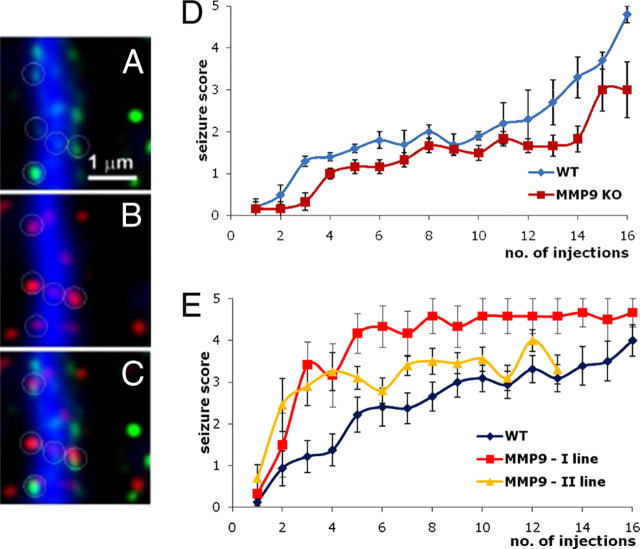
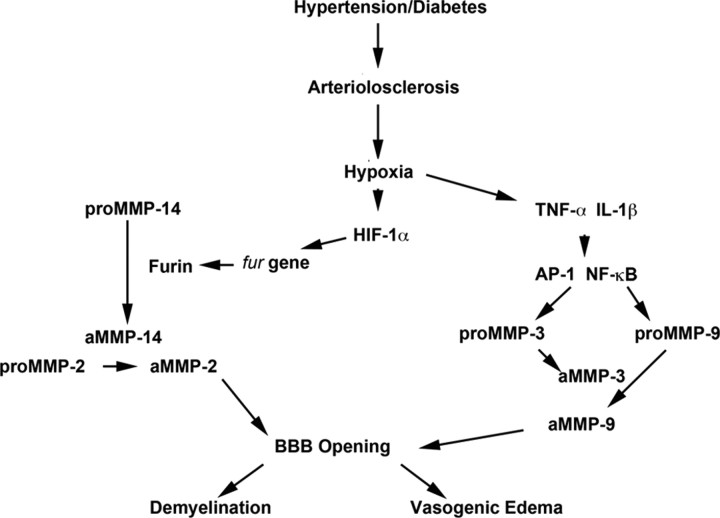

References
-
- Adair JC, Charlie J, Dencoff JE, Kaye JA, Quinn JF, Camicioli RM, Stetler-Stevenson WG, Rosenberg GA. Measurement of gelatinase B (MMP-9) in the cerebrospinal fluid of patients with vascular dementia and Alzheimer disease. Stroke. 2004;35:e159–e162. - PubMed
-
- Agrawal SM, Lau L, Yong VW. MMPs in the central nervous system: where the good guys go bad. Semin Cell Dev Biol. 2008;19:42–51. - PubMed
-
- Ahmed Z, Dent RG, Leadbeater WE, Smith C, Berry M, Logan A. Matrix metalloproteases: degradation of the inhibitory environment of the transected optic nerve and the scar by regenerating axons. Mol Cell Neurosci. 2005;28:64–78. - PubMed
-
- Ahmed Z, Mazibrada G, Seabright RJ, Dent RG, Berry M, Logan A. TACE-induced cleavage of NgR and p75NTR in dorsal root ganglion cultures disinhibits outgrowth and promotes branching of neurites in the presence of inhibitory CNS myelin. FASEB J. 2006;20:1939–1941. - PubMed
-
- Akiguchi I, Tomimoto H, Suenaga T, Wakita H, Budka H. Blood-brain barrier dysfunction in Binswanger's disease; an immunohistochemical study. Acta Neuropathol (Berl) 1998;95:78–84. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous