

Processing afferent proprioceptive information at the main cuneate nucleus of anesthetized cats
- PMID: 21084595
- PMCID: PMC6633671
- DOI: 10.1523/JNEUROSCI.2193-10.2010
Processing afferent proprioceptive information at the main cuneate nucleus of anesthetized cats
Abstract
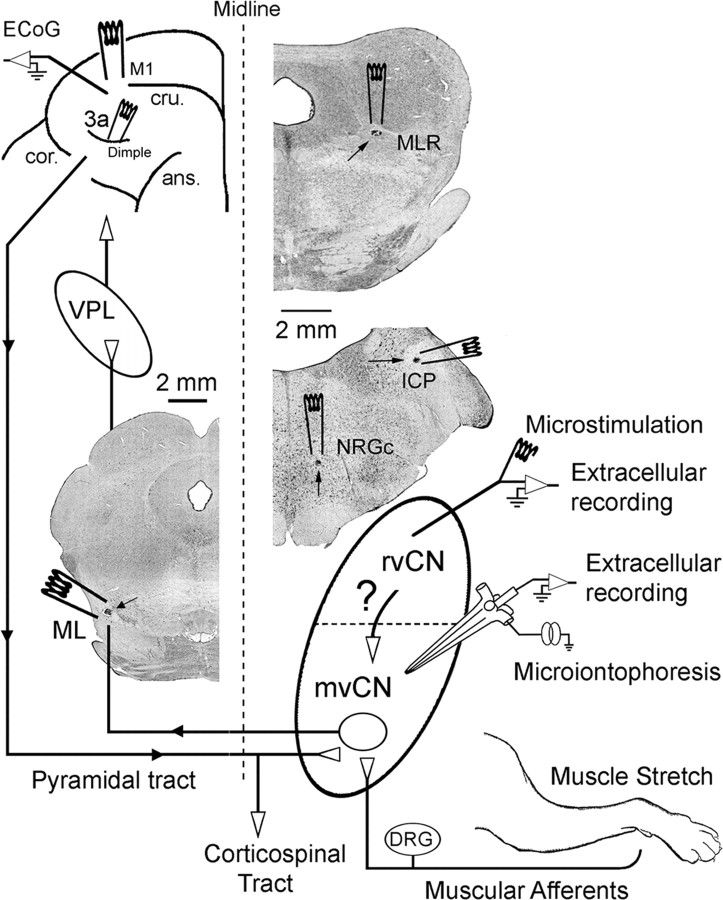
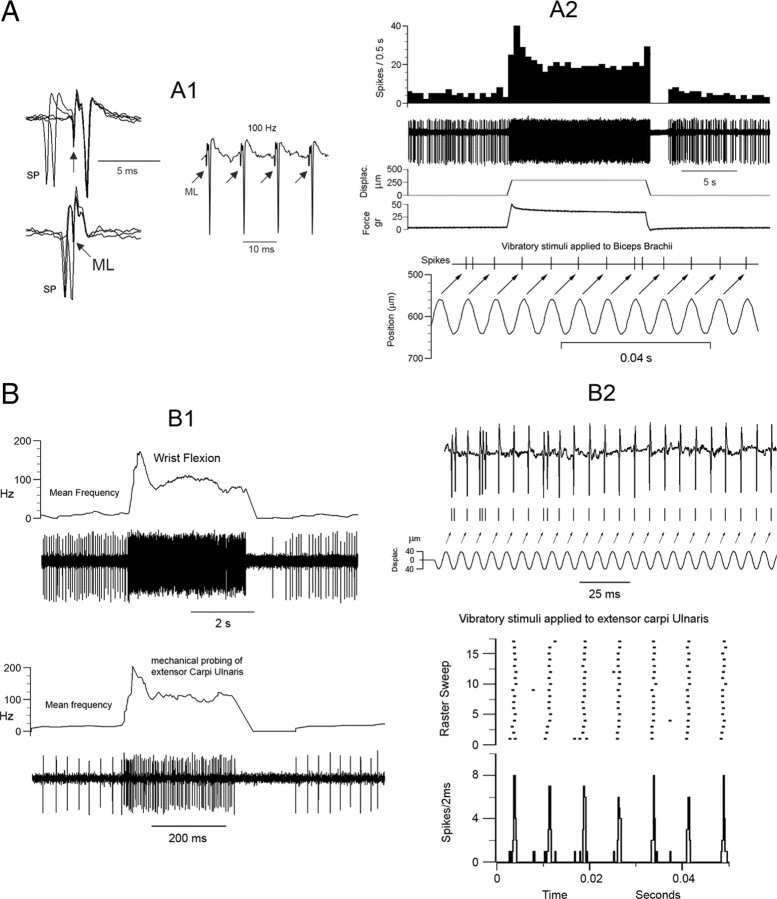
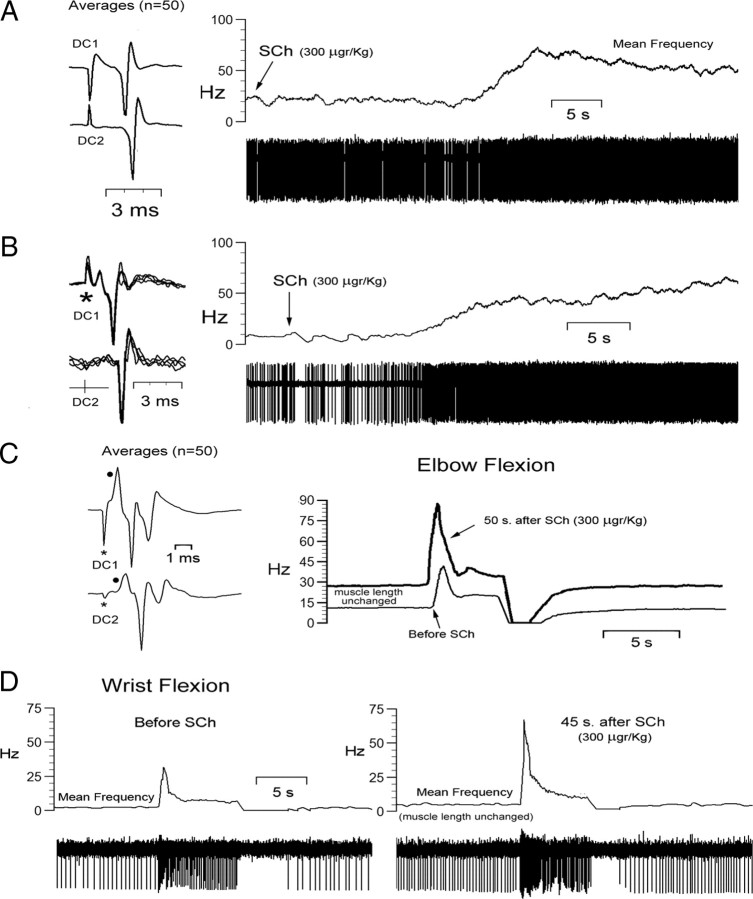
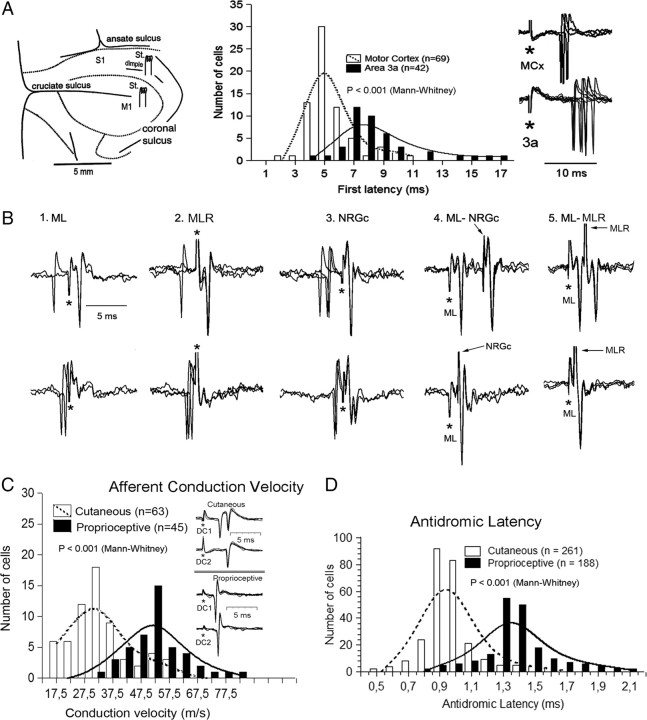
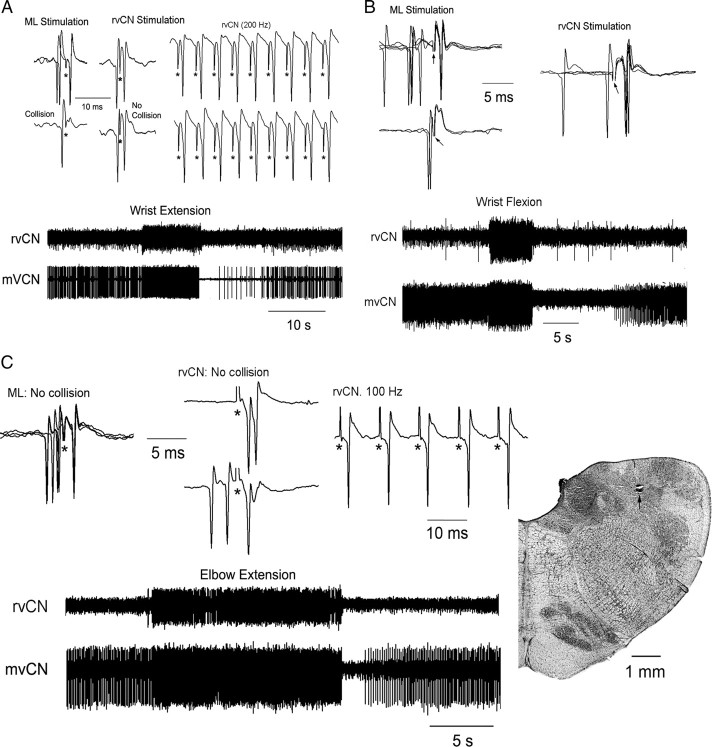
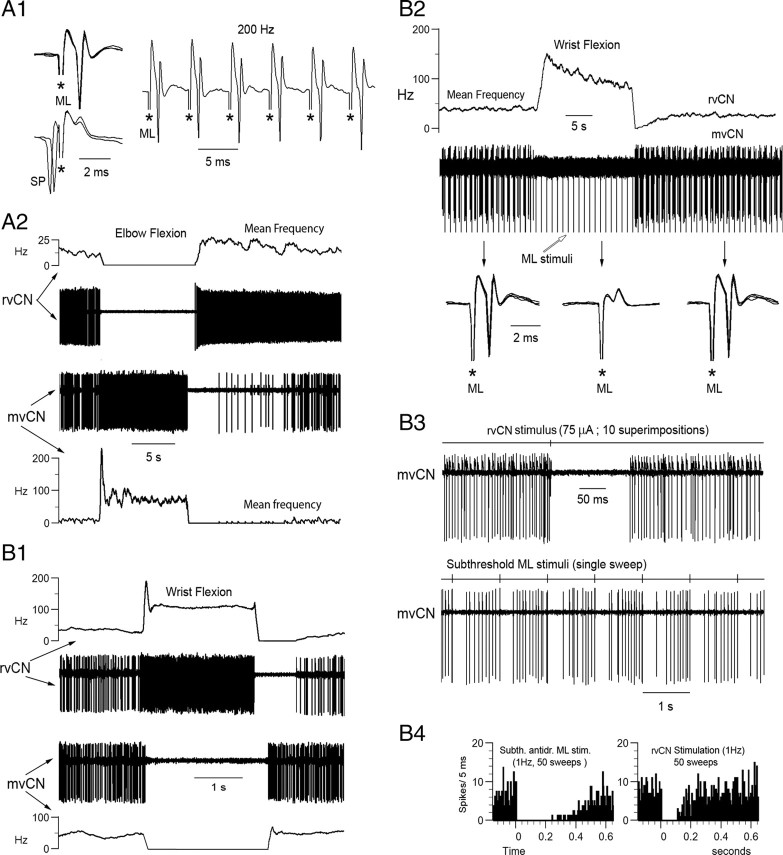
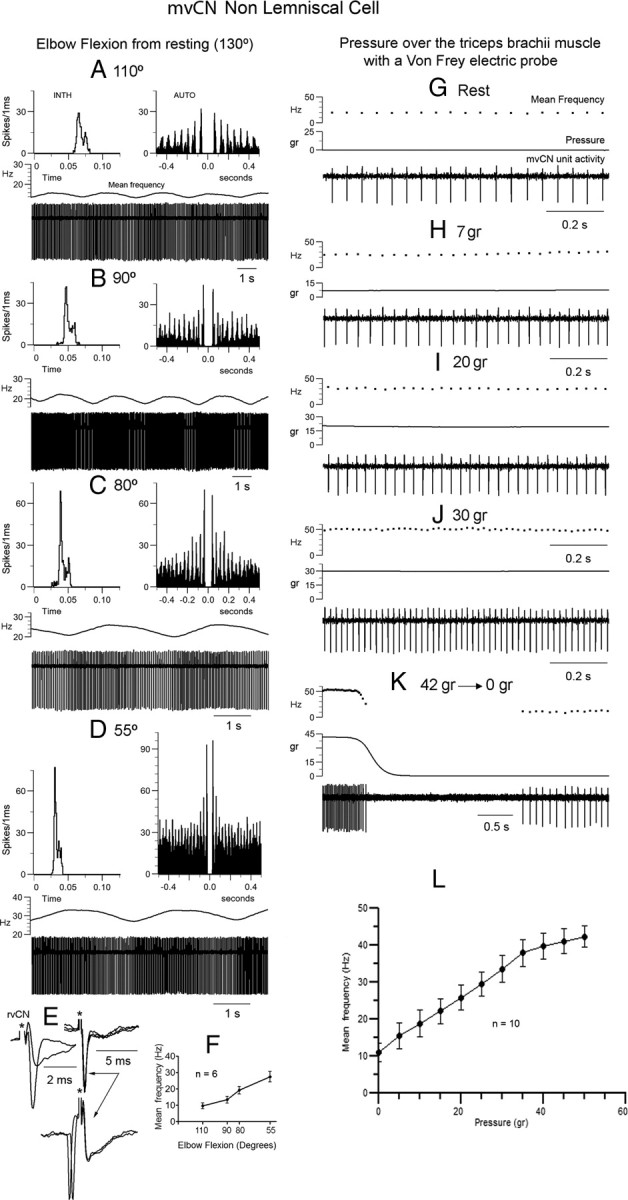
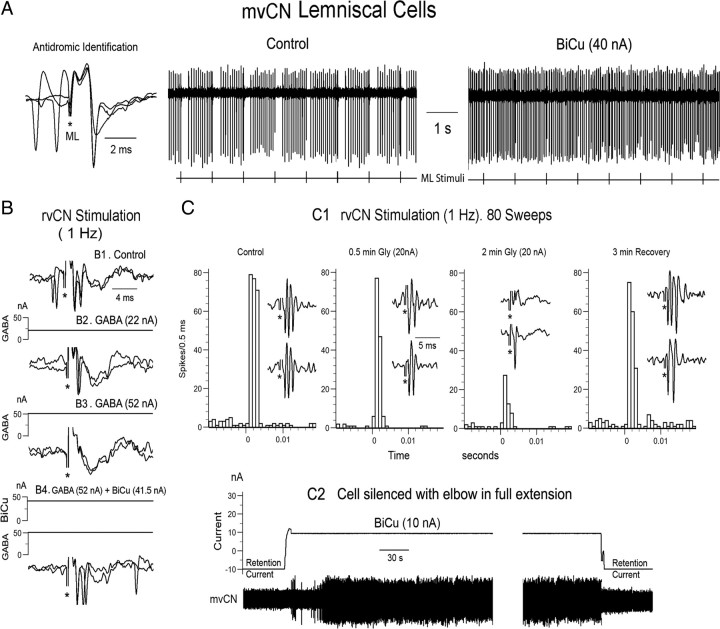
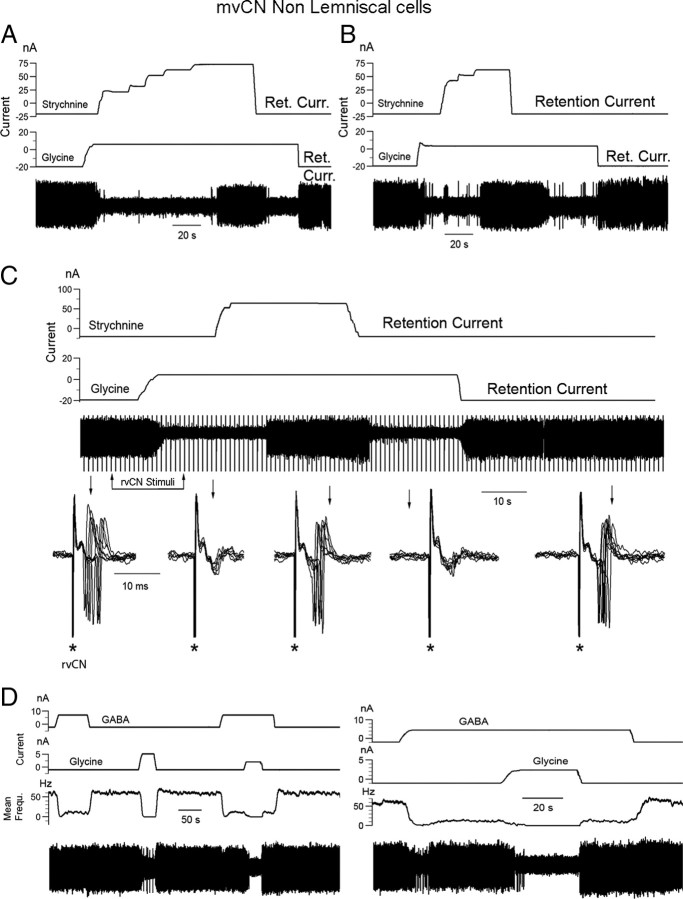
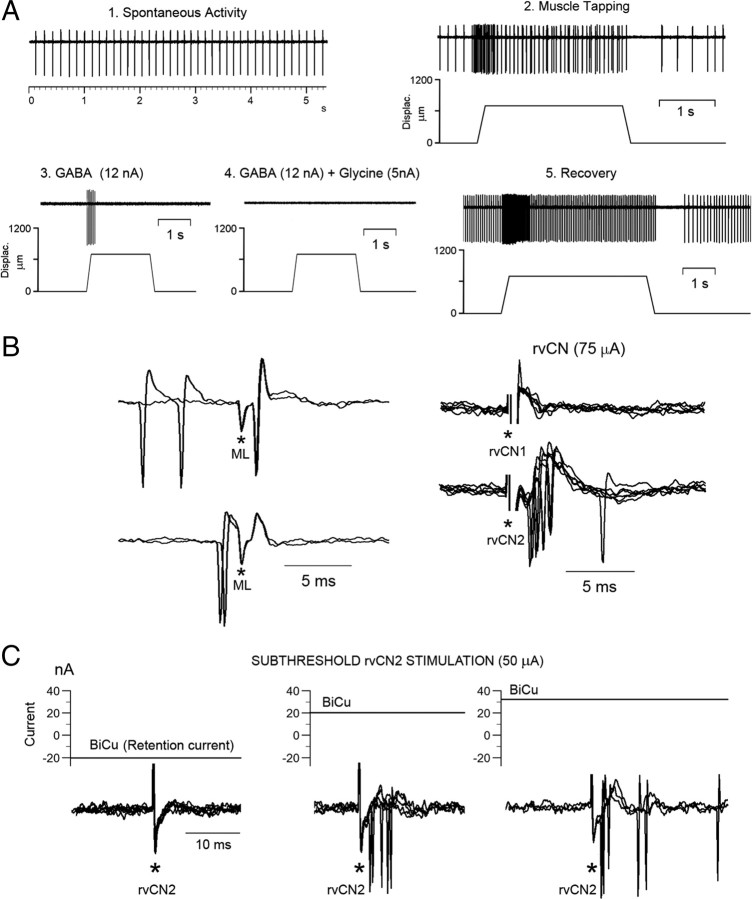
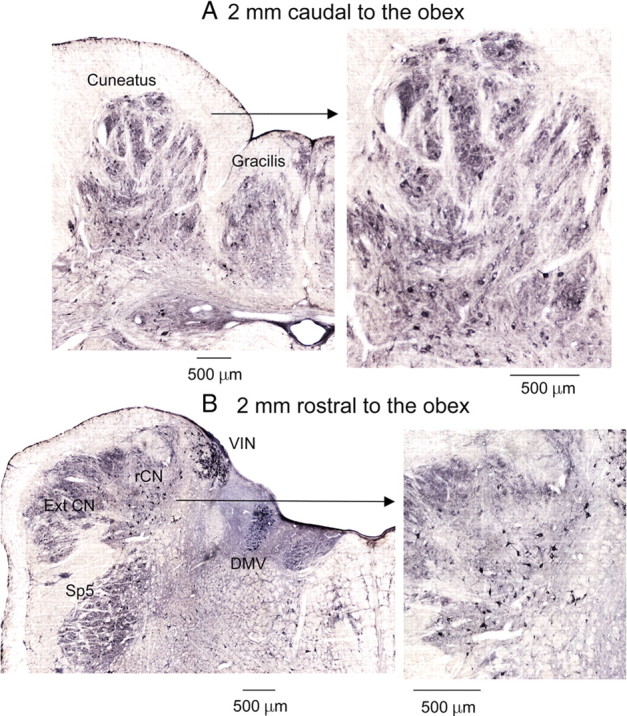
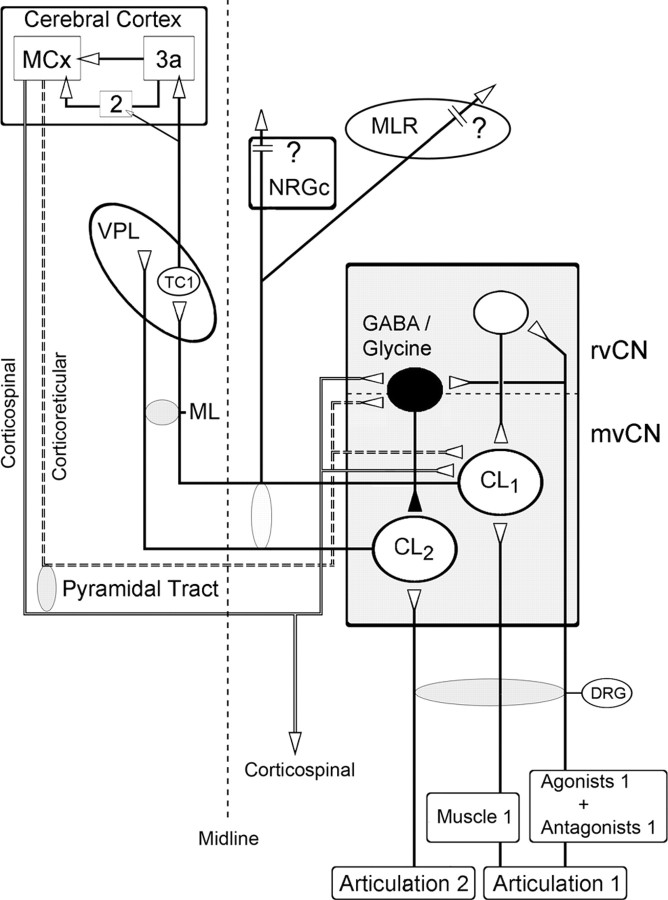
Medial lemniscal activity decreases before and during movement, suggesting prethalamic modulation, but the underlying mechanisms are largely unknown. Here we studied the mechanisms underlying proprioceptive transmission at the midventral cuneate nucleus (mvCN) of anesthetized cats using standard extracellular recordings combined with electrical stimulation and microiontophoresis. Dual simultaneous recordings from mvCN and rostroventral cuneate (rvCN) proprioceptive neurons demonstrated that microstimulation through the rvCN recording electrode induced dual effects on mvCN projection cells: potentiation when both neurons had excitatory receptive fields in muscles acting at the same joint, and inhibition when rvCN and mvCN cells had receptive fields located in different joints. GABA and/or glycine consistently abolished mvCN spontaneous and sensory-evoked activity, an effect reversed by bicuculline and strychnine, respectively; and immunohistochemistry data revealed that cells possessing strychnine-sensitive glycine receptors were uniformly distributed throughout the cuneate nucleus. It was also found that proprioceptive mvCN projection cells sent ipsilateral collaterals to the nucleus reticularis gigantocellularis and the mesencephalic locomotor region, and had slower antidromic conduction speeds than cutaneous fibers from the more dorsally located cluster region. The data suggest that (1) the rvCN-mvCM network is functionally related to joints rather than to single muscles producing an overall potentiation of proprioceptive feedback from a moving forelimb joint while inhibiting, through GABAergic and glycinergic interneurons, deep muscular feedback from other forelimb joints; and (2) mvCN projection cells collateralizing to or through the ipsilateral reticular formation allow for bilateral spreading of ascending proprioceptive feedback information.
Figures
Similar articles
-
Intracuneate mechanisms underlying primary afferent cutaneous processing in anaesthetized cats.Eur J Neurosci. 2004 Jun;19(11):3006-16. doi: 10.1111/j.0953-816X.2004.03432.x. Eur J Neurosci. 2004. PMID: 15182308
-
GABA(B) receptor-mediated modulation of cutaneous input at the cuneate nucleus in anesthetized cats.Neuroscience. 2006 Feb;137(3):1015-30. doi: 10.1016/j.neuroscience.2005.09.026. Epub 2005 Nov 17. Neuroscience. 2006. PMID: 16298083
-
Modulation and transmission of peripheral inputs in monkey cuneate and external cuneate nuclei.J Neurophysiol. 2011 Nov;106(5):2764-75. doi: 10.1152/jn.00449.2011. Epub 2011 Aug 24. J Neurophysiol. 2011. PMID: 21865437 Free PMC article.
-
Properties of proprioceptive neurons in the cuneate nucleus of the cat.J Neurophysiol. 1987 Apr;57(4):938-61. doi: 10.1152/jn.1987.57.4.938. J Neurophysiol. 1987. PMID: 3585464
-
The projection of joint receptors to the cuneate nucleus in the cat.J Physiol. 1980 Aug;305:433-49. doi: 10.1113/jphysiol.1980.sp013374. J Physiol. 1980. PMID: 7441563 Free PMC article.
Cited by
-
Pairing Transcranial Magnetic Stimulation and Loud Sounds Produces Plastic Changes in Motor Output.J Neurosci. 2023 Apr 5;43(14):2469-2481. doi: 10.1523/JNEUROSCI.0228-21.2022. Epub 2023 Mar 1. J Neurosci. 2023. PMID: 36859307 Free PMC article.
-
Proprioception: A New Era Set in Motion by Emerging Genetic and Bionic Strategies?Annu Rev Physiol. 2023 Feb 10;85:1-24. doi: 10.1146/annurev-physiol-040122-081302. Epub 2022 Nov 18. Annu Rev Physiol. 2023. PMID: 36400128 Free PMC article. Review.
-
Classification of Neurons in the Primate Reticular Formation and Changes after Recovery from Pyramidal Tract Lesion.J Neurosci. 2018 Jul 4;38(27):6190-6206. doi: 10.1523/JNEUROSCI.3371-17.2018. Epub 2018 May 23. J Neurosci. 2018. PMID: 29793974 Free PMC article.
-
Proprioception and the predictive sensing of active self-motion.Curr Opin Physiol. 2021 Apr;20:29-38. doi: 10.1016/j.cophys.2020.12.001. Epub 2021 Jan 22. Curr Opin Physiol. 2021. PMID: 33954270 Free PMC article.
-
Converging integration between ascending proprioceptive inputs and the corticospinal tract motor circuit underlying skilled movement control.Curr Opin Physiol. 2021 Feb;19:187-193. doi: 10.1016/j.cophys.2020.10.007. Epub 2020 Nov 13. Curr Opin Physiol. 2021. PMID: 33718693 Free PMC article.
References
-
- Aguilar J, Soto C, Rivadulla C, Canedo A. The lemniscal-cuneate recurrent excitation is suppressed by strychnine and enhanced by GABAa antagonists in the anaesthetized cat. Eur J Neurosci. 2002;16:1697–1704. - PubMed
-
- Aguilar J, Rivadulla C, Soto C, Canedo A. New corticocuneate cellular mechanisms underlying the modulation of cutaneous ascending transmission in anesthetized cats. J Neurophysiol. 2003;89:3328–3339. - PubMed
-
- Andersen P, Eccles JC, Schmidt RF, Yokota T. Slow potential waves produced in the cuneate nucleus by cutaneous volleys and by cortical stimulation. J Neurophysiol. 1964a;27:78–91. - PubMed
-
- Andersen P, Eccles JC, Oshima T, Schmidt RF. Mechanisms of synaptic transmission in the cuneate nucleus. J Neurophysiol. 1964b;27:1096–1116. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous