

Superfamily-wide portrait of serine hydrolase inhibition achieved by library-versus-library screening
- PMID: 21084632
- PMCID: PMC3000285
- DOI: 10.1073/pnas.1011663107
Superfamily-wide portrait of serine hydrolase inhibition achieved by library-versus-library screening
Abstract
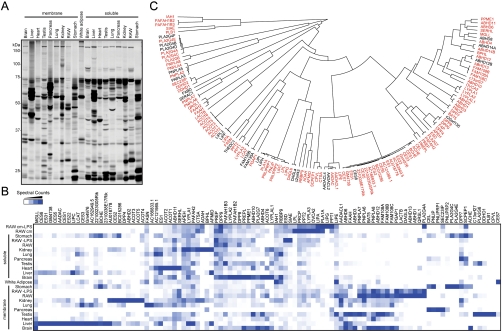
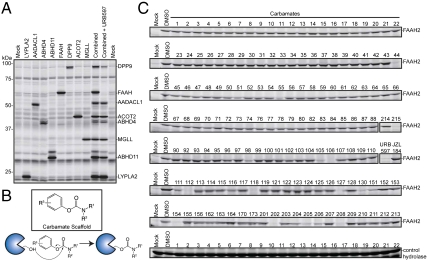
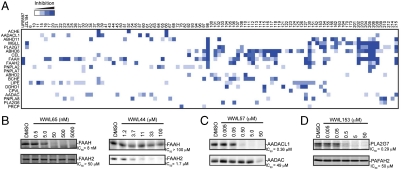
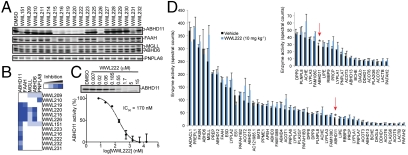
Serine hydrolases (SHs) are one of the largest and most diverse enzyme classes in mammals. They play fundamental roles in virtually all physiological processes and are targeted by drugs to treat diseases such as diabetes, obesity, and neurodegenerative disorders. Despite this, we lack biological understanding for most of the 110+ predicted mammalian metabolic SHs, in large part because of a dearth of assays to assess their biochemical activities and a lack of selective inhibitors to probe their function in living systems. We show here that the vast majority (> 80%) of mammalian metabolic SHs can be labeled in proteomes by a single, active site-directed fluorophosphonate probe. We exploit this universal activity-based assay in a library-versus-library format to screen 70+ SHs against 140+ structurally diverse carbamates. Lead inhibitors were discovered for ∼40% of the screened enzymes, including many poorly characterized SHs. Global profiles identified carbamate inhibitors that discriminate among highly sequence-related SHs and, conversely, enzymes that share inhibitor sensitivity profiles despite lacking sequence homology. These findings indicate that sequence relatedness is not a strong predictor of shared pharmacology within the SH superfamily. Finally, we show that lead carbamate inhibitors can be optimized into pharmacological probes that inactivate individual SHs with high specificity in vivo.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Gerlt JA, Babbitt PC. Divergent evolution of enzymatic function: Mechanistically diverse superfamilies and functionally distinct suprafamilies. Annu Rev Biochem. 2001;70:209–246. - PubMed
-
- Xiao SH, et al. High throughput screening for orphan and liganded GPCRs. Comb Chem High Throughput Screening. 2008;11(3):195–215. - PubMed
-
- Fabian MA, et al. A small molecule-kinase interaction map for clinical kinase inhibitors. Nat Biotechnol. 2005;23(3):329–336. - PubMed
-
- Bishop AC, et al. A chemical switch for inhibitor-sensitive alleles of any protein kinase. Nature. 2000;407:395–401. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
