

Sumoylation activates the transcriptional activity of Pax-6, an important transcription factor for eye and brain development
- PMID: 21084637
- PMCID: PMC3000302
- DOI: 10.1073/pnas.1007866107
Sumoylation activates the transcriptional activity of Pax-6, an important transcription factor for eye and brain development
Abstract
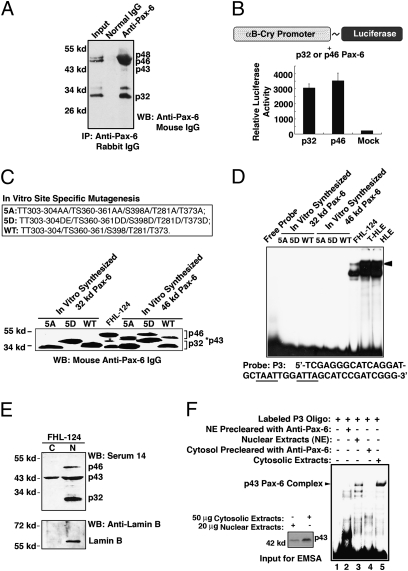
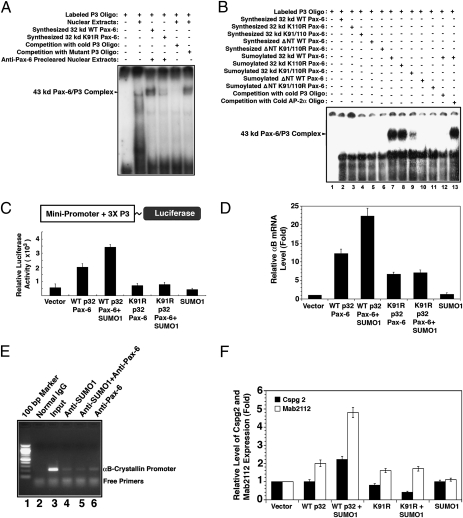
Pax-6 is an evolutionarily conserved transcription factor regulating brain and eye development. Four Pax-6 isoforms have been reported previously. Although the longer Pax-6 isoforms (p46 and p48) bear two DNA-binding domains, the paired domain (PD) and the homeodomain (HD), the shorter Pax-6 isoform p32 contains only the HD for DNA binding. Although a third domain, the proline-, serine- and threonine-enriched activation (PST) domain, in the C termini of all Pax-6 isoforms mediates their transcriptional modulation via phosphorylation, how p32 Pax-6 could regulate target genes remains to be elucidated. In the present study, we show that sumoylation at K91 is required for p32 Pax-6 to bind to a HD-specific site and regulate expression of target genes. First, in vitro-synthesized p32 Pax-6 alone cannot bind the P3 sequence, which contains the HD recognition site, unless it is preincubated with nuclear extracts precleared by anti-Pax-6 but not by anti-small ubiquitin-related modifier 1 (anti-SUMO1) antibody. Second, in vitro-synthesized p32 Pax-6 can be sumoylated by SUMO1, and the sumoylated p32 Pax-6 then can bind to the P3 sequence. Third, Pax-6 and SUMO1 are colocalized in the embryonic optic and lens vesicles and can be coimmunoprecipitated. Finally, SUMO1-conjugated p32 Pax-6 exists in both the nucleus and cytoplasm, and sumoylation significantly enhances the DNA-binding ability of p32 Pax-6 and positively regulates gene expression. Together, our results demonstrate that sumoylation activates p32 Pax-6 in both DNA-binding and transcriptional activities. In addition, our studies demonstrate that p32 and p46 Pax-6 possess differential DNA-binding and regulatory activities.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


References
-
- Mansouri A, Hallonet M, Gruss P. Pax genes and their roles in cell differentiation and development. Curr Opin Cell Biol. 1996;8:851–857. - PubMed
-
- Callaerts P, Halder G, Gehring WJ. PAX-6 in development and evolution. Annu Rev Neurosci. 1997;20:483–532. - PubMed
-
- Ton CC, et al. Positional cloning and characterization of a paired box- and homeobox-containing gene from the aniridia region. Cell. 1991;67:1059–1074. - PubMed
-
- Glaser T, Walton DS, Maas RL. Genomic structure, evolutionary conservation and aniridia mutations in the human PAX6 gene. Nat Genet. 1992;2:232–239. - PubMed
-
- Wilson D, Sheng G, Lecuit T, Dostatni N, Desplan C. Cooperative dimerization of paired class homeo domains on DNA. Genes Dev. 1993;7:2120–2134. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
