

Formation, regulation and evolution of Caenorhabditis elegans 3'UTRs
- PMID: 21085120
- PMCID: PMC3057491
- DOI: 10.1038/nature09616
Formation, regulation and evolution of Caenorhabditis elegans 3'UTRs
Abstract
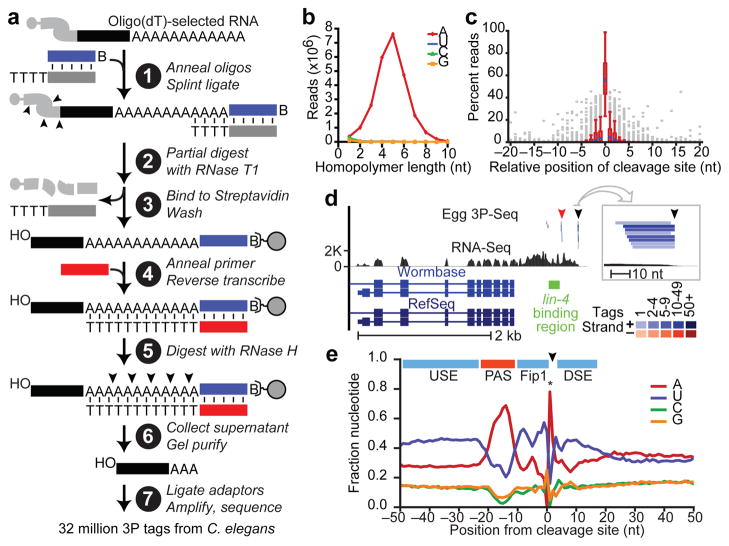
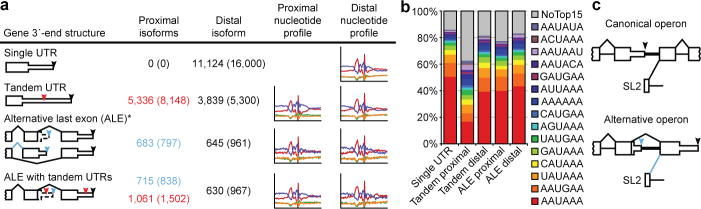
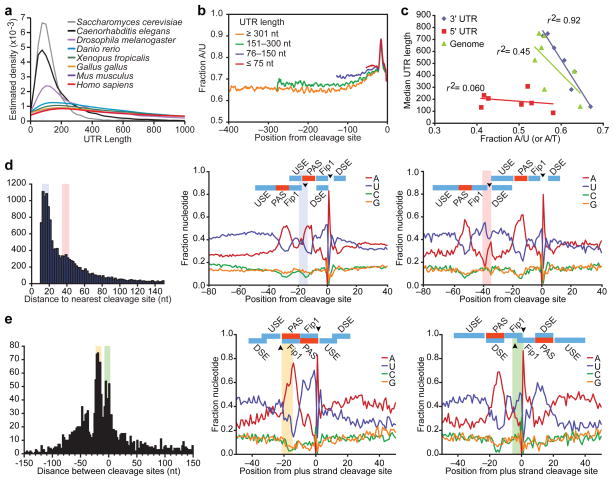
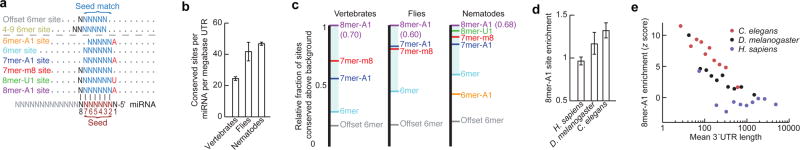
Post-transcriptional gene regulation frequently occurs through elements in mRNA 3' untranslated regions (UTRs). Although crucial roles for 3'UTR-mediated gene regulation have been found in Caenorhabditis elegans, most C. elegans genes have lacked annotated 3'UTRs. Here we describe a high-throughput method for reliable identification of polyadenylated RNA termini, and we apply this method, called poly(A)-position profiling by sequencing (3P-Seq), to determine C. elegans 3'UTRs. Compared to standard methods also recently applied to C. elegans UTRs, 3P-Seq identified 8,580 additional UTRs while excluding thousands of shorter UTR isoforms that do not seem to be authentic. Analysis of this expanded and corrected data set suggested that the high A/U content of C. elegans 3'UTRs facilitated genome compaction, because the elements specifying cleavage and polyadenylation, which are A/U rich, can more readily emerge in A/U-rich regions. Indeed, 30% of the protein-coding genes have mRNAs with alternative, partially overlapping end regions that generate another 10,480 cleavage and polyadenylation sites that had gone largely unnoticed and represent potential evolutionary intermediates of progressive UTR shortening. Moreover, a third of the convergently transcribed genes use palindromic arrangements of bidirectional elements to specify UTRs with convergent overlap, which also contributes to genome compaction by eliminating regions between genes. Although nematode 3'UTRs have median length only one-sixth that of mammalian 3'UTRs, they have twice the density of conserved microRNA sites, in part because additional types of seed-complementary sites are preferentially conserved. These findings reveal the influence of cleavage and polyadenylation on the evolution of genome architecture and provide resources for studying post-transcriptional gene regulation.
Figures
References
-
- Moore MJ. From birth to death: the complex lives of eukaryotic mRNAs. Science. 2005;309:1514–1518. - PubMed
-
- Ahringer J, Kimble J. Control of the sperm-oocyte switch in Caenorhabditis elegans hermaphrodites by the fem-3 3' untranslated region. Nature. 1991;349:346–348. - PubMed
-
- Wightman B, Burglin TR, Gatto J, Arasu P, Ruvkun G. Negative regulatory sequences in the lin-14 3'-untranslated region are necessary to generate a temporal switch during Caenorhabditis elegans development. Genes Dev. 1991;5:1813–1824. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
