

Staphylococcus aureus hijacks a skin commensal to intensify its virulence: immunization targeting β-hemolysin and CAMP factor
- PMID: 21085191
- PMCID: PMC3057116
- DOI: 10.1038/jid.2010.319
Staphylococcus aureus hijacks a skin commensal to intensify its virulence: immunization targeting β-hemolysin and CAMP factor
Abstract
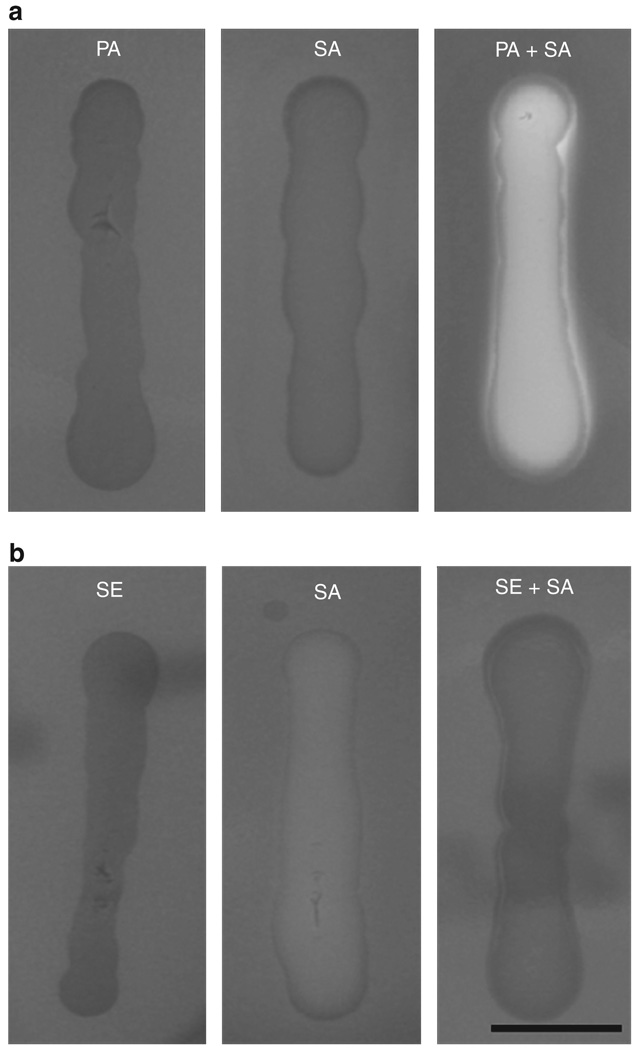
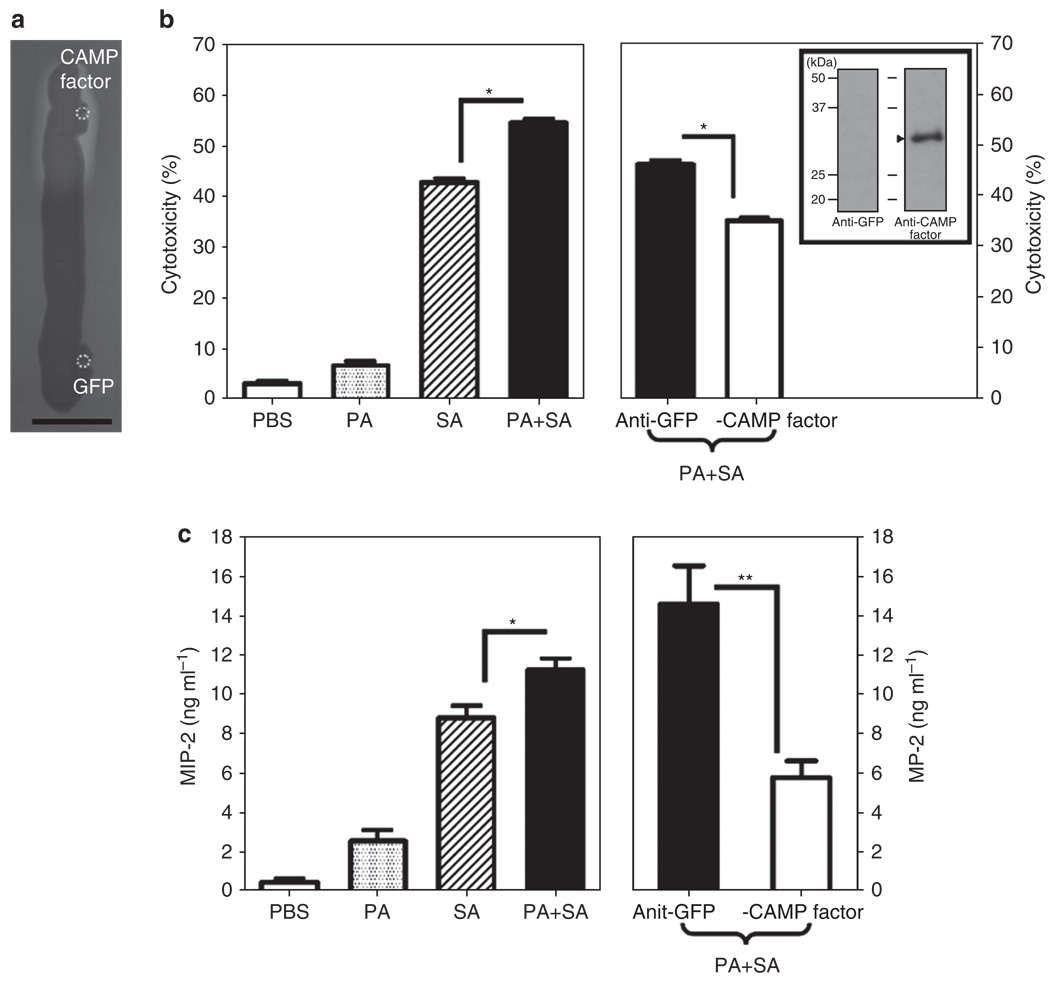
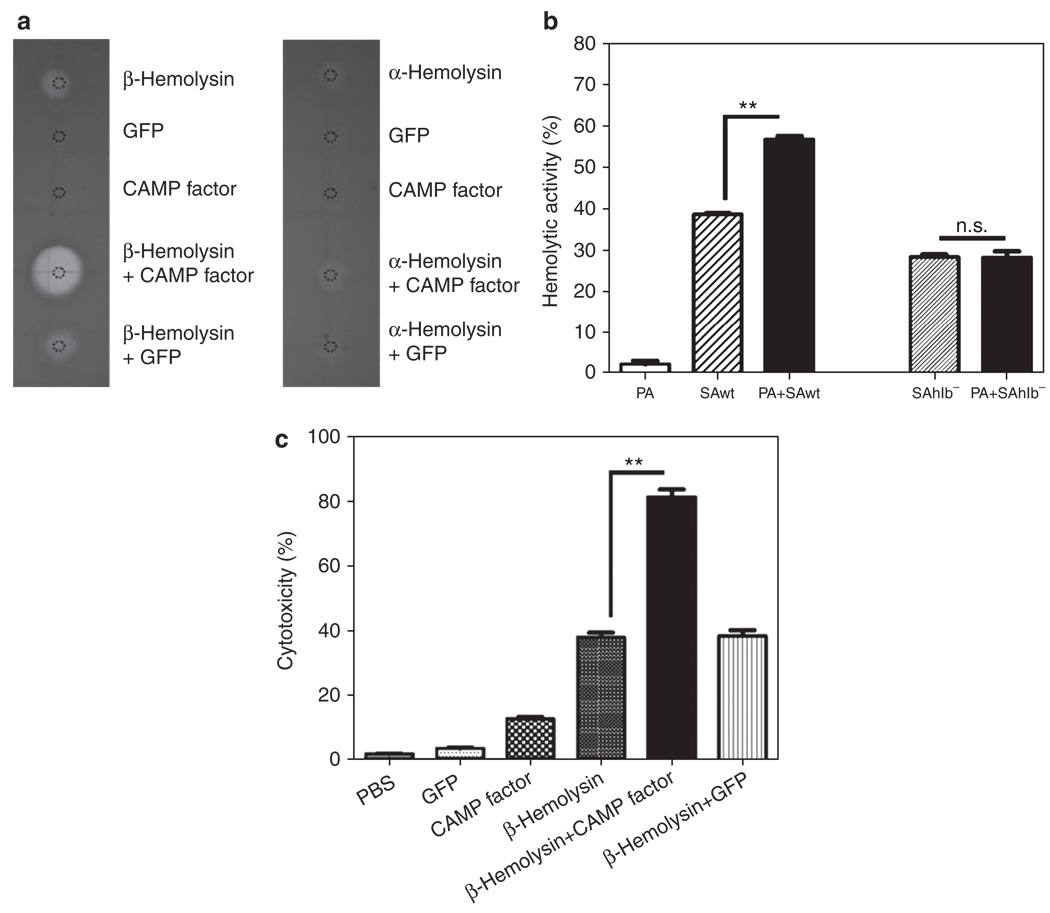
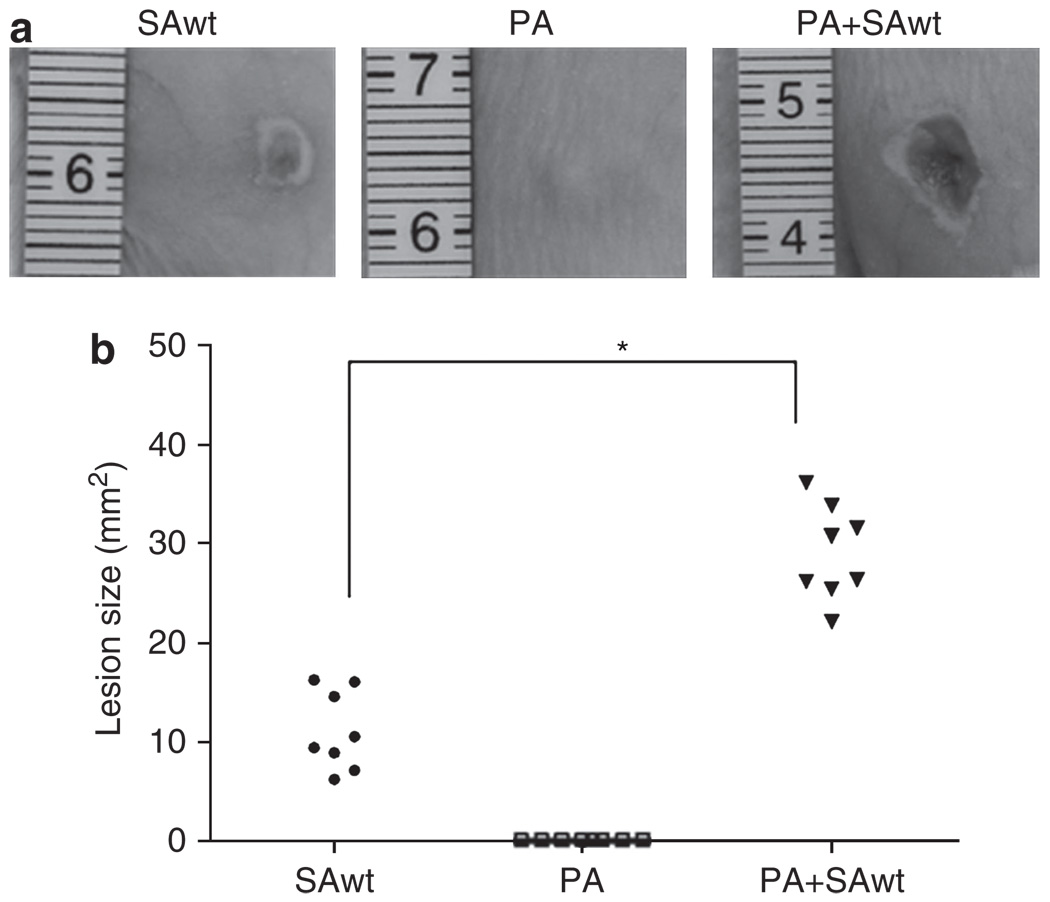
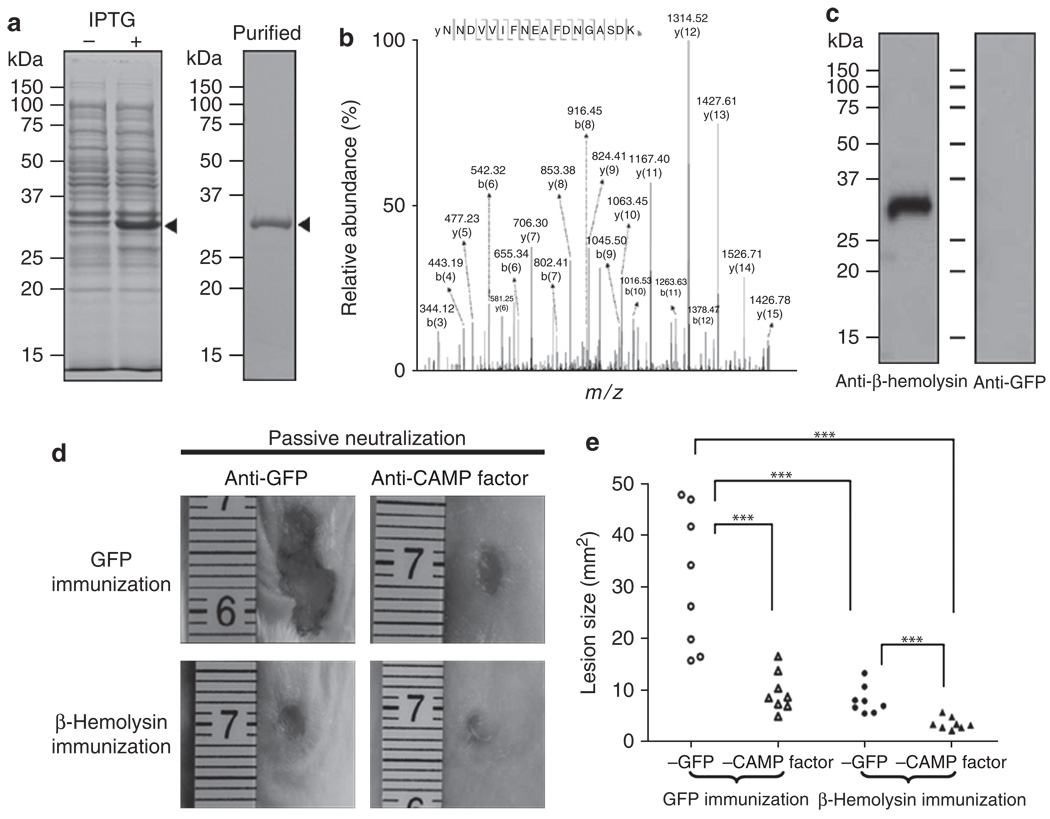
The need for a new anti-Staphylococcus aureus therapy that can effectively cripple bacterial infection, neutralize secretory virulence factors, and lower the risk of creating bacterial resistance is undisputed. Here, we propose what is, to our knowledge, a previously unreported infectious mechanism by which S. aureus may commandeer Propionibacterium acnes, a key member of the human skin microbiome, to spread its invasion and highlight two secretory virulence factors (S. aureus β-hemolysin and P. acnes CAMP (Christie, Atkins, Munch-Peterson) factor) as potential molecular targets for immunotherapy against S. aureus infection. Our data demonstrate that the hemolysis and cytolysis by S. aureus were noticeably augmented when S. aureus was grown with P. acnes. The augmentation was significantly abrogated when the P. acnes CAMP factor was neutralized or β-hemolysin of S. aureus was mutated. In addition, the hemolysis and cytolysis of recombinant β-hemolysin were markedly enhanced by recombinant CAMP factor. Furthermore, P. acnes exacerbated S. aureus-induced skin lesions in vivo. The combination of CAMP factor neutralization and β-hemolysin immunization cooperatively suppressed the skin lesions caused by coinfection of P. acnes and S. aureus. These observations suggest a previously unreported immunotherapy targeting the interaction of S. aureus with a skin commensal.
Conflict of interest statement
The authors state no conflict of interest.
Figures
Similar articles
-
Passive immunoprotection targeting a secreted CAMP factor of Propionibacterium acnes as a novel immunotherapeutic for acne vulgaris.Vaccine. 2011 Apr 12;29(17):3230-8. doi: 10.1016/j.vaccine.2011.02.036. Epub 2011 Feb 26. Vaccine. 2011. PMID: 21354482 Free PMC article.
-
The Anti-Inflammatory Activities of Propionibacterium acnes CAMP Factor-Targeted Acne Vaccines.J Invest Dermatol. 2018 Nov;138(11):2355-2364. doi: 10.1016/j.jid.2018.05.032. Epub 2018 Jun 30. J Invest Dermatol. 2018. PMID: 29964032
-
Propionibacterium acnes CAMP factor and host acid sphingomyelinase contribute to bacterial virulence: potential targets for inflammatory acne treatment.PLoS One. 2011 Apr 12;6(4):e14797. doi: 10.1371/journal.pone.0014797. PLoS One. 2011. PMID: 21533261 Free PMC article.
-
Prospects of acne vaccines targeting secreted virulence factors of Cutibacterium acnes.Expert Rev Vaccines. 2019 May;18(5):433-437. doi: 10.1080/14760584.2019.1593830. Epub 2019 Mar 28. Expert Rev Vaccines. 2019. PMID: 30920859 Review.
-
The potential use of toxin antibodies as a strategy for controlling acute Staphylococcus aureus infections.Expert Opin Ther Targets. 2012 Jun;16(6):601-12. doi: 10.1517/14728222.2012.682573. Epub 2012 Apr 25. Expert Opin Ther Targets. 2012. PMID: 22530584 Free PMC article. Review.
Cited by
-
Passive immunoprotection targeting a secreted CAMP factor of Propionibacterium acnes as a novel immunotherapeutic for acne vulgaris.Vaccine. 2011 Apr 12;29(17):3230-8. doi: 10.1016/j.vaccine.2011.02.036. Epub 2011 Feb 26. Vaccine. 2011. PMID: 21354482 Free PMC article.
-
Group B Streptococcus CAMP Factor Does Not Contribute to Interactions with the Vaginal Epithelium and Is Dispensable for Vaginal Colonization in Mice.Microbiol Spectr. 2021 Dec 22;9(3):e0105821. doi: 10.1128/Spectrum.01058-21. Epub 2021 Dec 15. Microbiol Spectr. 2021. PMID: 34908468 Free PMC article.
-
Changes in the Microbiome During Chronic Rhinosinusitis.Pathogens. 2024 Dec 30;14(1):14. doi: 10.3390/pathogens14010014. Pathogens. 2024. PMID: 39860975 Free PMC article. Review.
-
Insights into microbial dysbiosis and Cutibacterium acnes CAMP factor interactions in acne vulgaris.Microb Genom. 2025 Jul;11(7):001449. doi: 10.1099/mgen.0.001449. Microb Genom. 2025. PMID: 40668089 Free PMC article.
-
Efficacy of Bacteriophages in Propionibacterium acnes-Induced Inflammation in Mice.Ann Dermatol. 2019 Feb;31(1):22-28. doi: 10.5021/ad.2019.31.1.22. Epub 2019 Jan 2. Ann Dermatol. 2019. PMID: 33911535 Free PMC article.
References
-
- Ahn CY, Ko CY, Wagar EA, et al. Microbial evaluation: 139 implants removed from symptomatic patients. Plast Reconstr Surg. 1996;98:1225–1229. - PubMed
-
- Bashir A, Mujahid TY, Jehan N. Antibiotic resistance profile: isolation and characterization of clinical isolates of staphylococci from patients with community-acquired skin infections. Pak J Pharm Sci. 2007;20:299–304. - PubMed
-
- Brook I, Frazier EH. Infections caused by Propionibacterium species. Rev Infect Dis. 1991;13:819–822. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
