

Host cell invasion and virulence mediated by Candida albicans Ssa1
- PMID: 21085601
- PMCID: PMC2978716
- DOI: 10.1371/journal.ppat.1001181
Host cell invasion and virulence mediated by Candida albicans Ssa1
Abstract
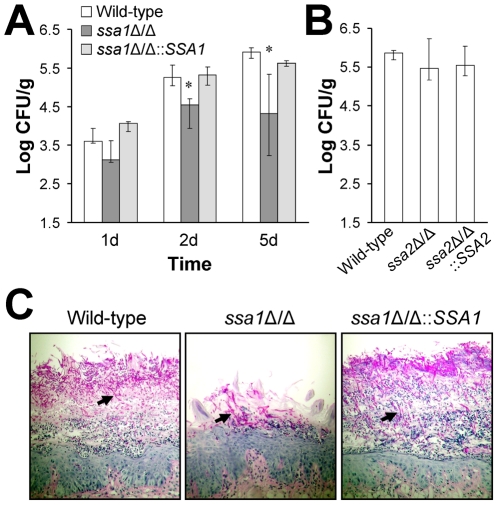
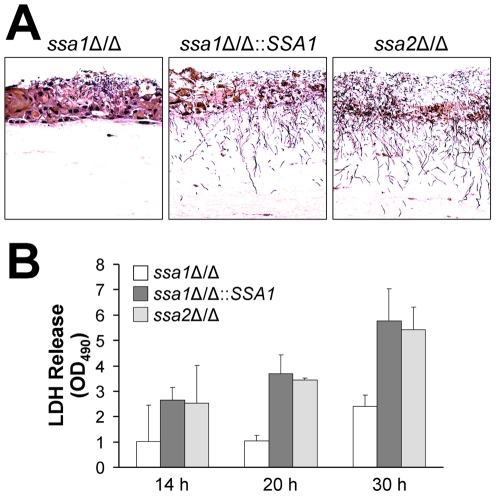
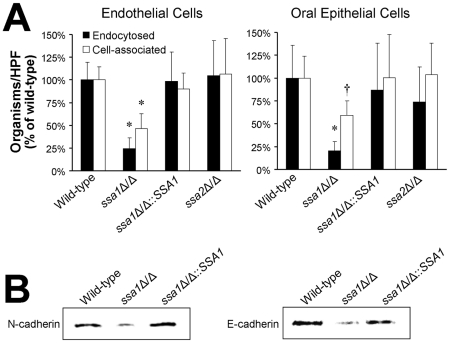
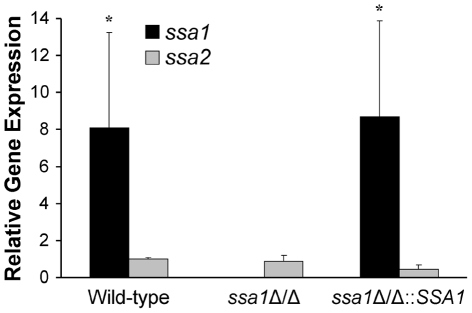
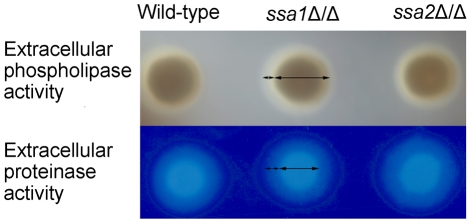
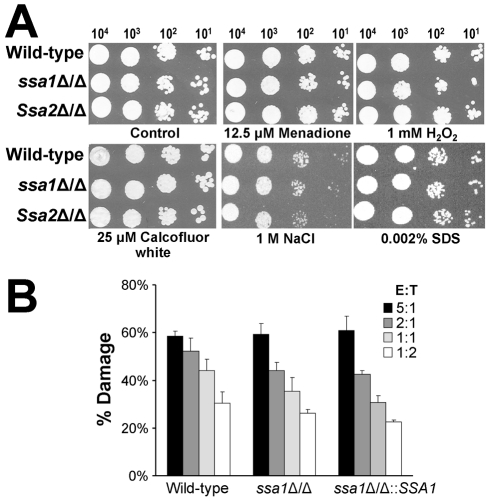
Candida albicans Ssa1 and Ssa2 are members of the HSP70 family of heat shock proteins that are expressed on the cell surface and function as receptors for antimicrobial peptides such as histatins. We investigated the role of Ssa1 and Ssa2 in mediating pathogenic host cell interactions and virulence. A C. albicans ssa1Δ/Δ mutant had attenuated virulence in murine models of disseminated and oropharyngeal candidiasis, whereas an ssa2Δ/Δ mutant did not. In vitro studies revealed that the ssa1Δ/Δ mutant caused markedly less damage to endothelial cells and oral epithelial cell lines. Also, the ssa1Δ/Δ mutant had defective binding to endothelial cell N-cadherin and epithelial cell E-cadherin, receptors that mediate host cell endocytosis of C. albicans. As a result, this mutant had impaired capacity to induce its own endocytosis by endothelial cells and oral epithelial cells. Latex beads coated with recombinant Ssa1 were avidly endocytosed by both endothelial cells and oral epithelial cells, demonstrating that Ssa1 is sufficient to induce host cell endocytosis. These results indicate that Ssa1 is a novel invasin that binds to host cell cadherins, induces host cell endocytosis, and is critical for C. albicans to cause maximal damage to host cells and induce disseminated and oropharyngeal disease.
Conflict of interest statement
SGF holds equity in NovaDigm Therapeutics.
Figures





References
-
- Wisplinghoff H, Bischoff T, Tallent SM, Seifert H, Wenzel RP, et al. Nosocomial bloodstream infections in US hospitals: analysis of 24,179 cases from a prospective nationwide surveillance study. Clin Infect Dis. 2004;39:309–317. - PubMed
-
- Filler SG. Candida-host cell receptor-ligand interactions. Curr Opin Microbiol. 2006;9:333–339. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
