

Transient increase in cyclic AMP localized to macrophage phagosomes
- PMID: 21085604
- PMCID: PMC2978719
- DOI: 10.1371/journal.pone.0013962
Transient increase in cyclic AMP localized to macrophage phagosomes
Abstract
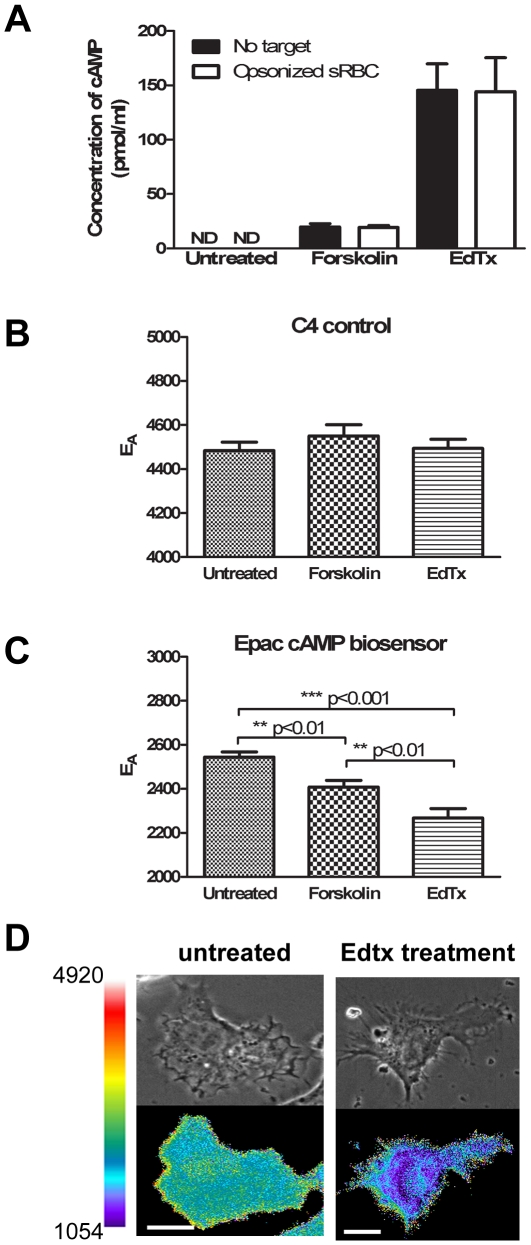
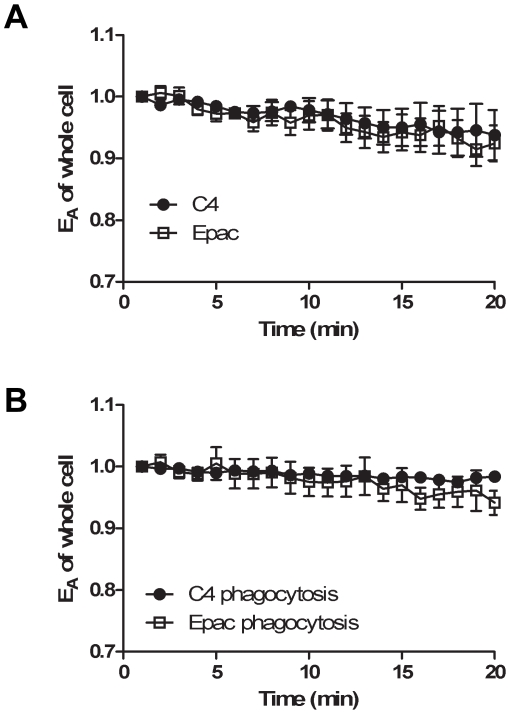
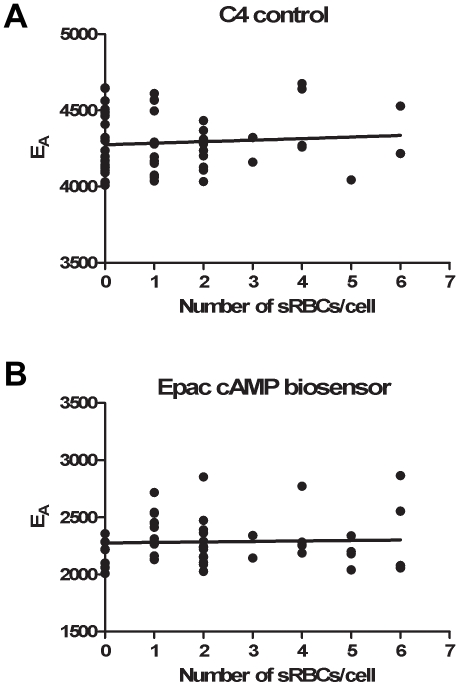
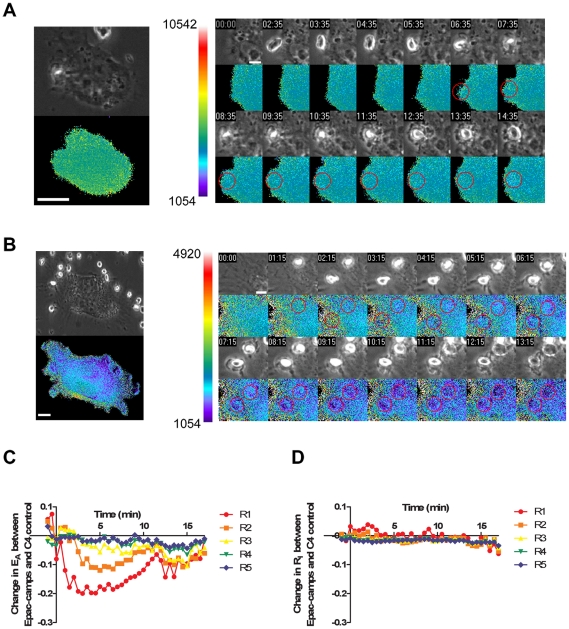
Cyclic AMP (cAMP) regulates many biological processes and cellular functions. The importance of spatially localized intracellular gradients of cAMP is increasingly appreciated. Previous work in macrophages has shown that cAMP is produced during phagocytosis and that elevated cAMP levels suppress host defense functions, including generation of proinflammatory mediators, phagocytosis and killing. However, the spatial and kinetic characteristics of cAMP generation in phagocytosing macrophages have yet to be examined. Using a Förster resonance energy transfer (FRET)-based cAMP biosensor, we measured the generation of cAMP in live macrophages. We detected no difference in bulk intracellular cAMP levels between resting cells and cells actively phagocytosing IgG-opsonized particles. However, analysis with the biosensor revealed a rapid decrease in FRET signal corresponding to a transient burst of cAMP production localized to the forming phagosome. cAMP levels returned to baseline after the particle was internalized. These studies indicate that localized increases in cAMP accompany phagosome formation and provide a framework for a more complete understanding of how cAMP regulates macrophage host defense functions.
Conflict of interest statement
Figures
Similar articles
-
SNAP-23 regulates phagosome formation and maturation in macrophages.Mol Biol Cell. 2012 Dec;23(24):4849-63. doi: 10.1091/mbc.E12-01-0069. Epub 2012 Oct 19. Mol Biol Cell. 2012. PMID: 23087210 Free PMC article.
-
Compartmentalization of PDE-4 and cAMP-dependent protein kinase in neutrophils and macrophages during phagocytosis.Cell Biochem Biophys. 1998;28(2-3):251-75. doi: 10.1007/BF02737813. Cell Biochem Biophys. 1998. PMID: 9515168
-
Spectral imaging of FRET-based sensors reveals sustained cAMP gradients in three spatial dimensions.Cytometry A. 2018 Oct;93(10):1029-1038. doi: 10.1002/cyto.a.23572. Epub 2018 Sep 3. Cytometry A. 2018. PMID: 30176184 Free PMC article.
-
Decoding spatial and temporal features of neuronal cAMP/PKA signaling with FRET biosensors.Biotechnol J. 2014 Feb;9(2):192-202. doi: 10.1002/biot.201300202. Epub 2014 Jan 29. Biotechnol J. 2014. PMID: 24478276 Review.
-
cAMP/PKA signaling compartmentalization in cardiomyocytes: Lessons from FRET-based biosensors.J Mol Cell Cardiol. 2019 Jun;131:112-121. doi: 10.1016/j.yjmcc.2019.04.020. Epub 2019 Apr 24. J Mol Cell Cardiol. 2019. PMID: 31028775 Review.
Cited by
-
OR2AT4 and OR1A2 counterregulate molecular pathophysiological processes of steroid-resistant inflammatory lung diseases in human alveolar macrophages.Mol Med. 2022 Dec 12;28(1):150. doi: 10.1186/s10020-022-00572-8. Mol Med. 2022. PMID: 36503361 Free PMC article.
-
New DAG and cAMP Sensors Optimized for Live-Cell Assays in Automated Laboratories.J Biomol Screen. 2016 Mar;21(3):298-305. doi: 10.1177/1087057115618608. Epub 2015 Dec 11. J Biomol Screen. 2016. PMID: 26657040 Free PMC article.
-
The promising roles of macrophages in geriatric hip fracture.Front Cell Dev Biol. 2022 Aug 26;10:962990. doi: 10.3389/fcell.2022.962990. eCollection 2022. Front Cell Dev Biol. 2022. PMID: 36092716 Free PMC article. Review.
-
Leptin enhances availability of apoptotic cell-derived self-antigen in systemic lupus erythematosus.PLoS One. 2014 Nov 17;9(11):e112826. doi: 10.1371/journal.pone.0112826. eCollection 2014. PLoS One. 2014. PMID: 25401752 Free PMC article.
-
Macrophage TLR4 and PAR2 Signaling: Role in Regulating Vascular Inflammatory Injury and Repair.Front Immunol. 2020 Sep 18;11:2091. doi: 10.3389/fimmu.2020.02091. eCollection 2020. Front Immunol. 2020. PMID: 33072072 Free PMC article. Review.
References
-
- Swanson JA, Hoppe AD. The coordination of signaling during Fc receptor-mediated phagocytosis. J Leukoc Biol. 2004;76:1093–1103. - PubMed
-
- Medzhitov R. Approaching the asymptote: 20 years later. Immunity. 2009;30:766–775. - PubMed
-
- van Lookeren Campagne M, Wiesmann C, Brown EJ. Macrophage complement receptors and pathogen clearance. Cell Microbiol. 2007;9:2095–2102. - PubMed
-
- Forman HJ, Torres M. Reactive oxygen species and cell signaling: respiratory burst in macrophage signaling. Am J Respir Crit Care Med. 2002;166:S4–8. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
