

Quantitative control of organ shape by combinatorial gene activity
- PMID: 21085695
- PMCID: PMC2976723
- DOI: 10.1371/journal.pbio.1000538
Quantitative control of organ shape by combinatorial gene activity
Abstract
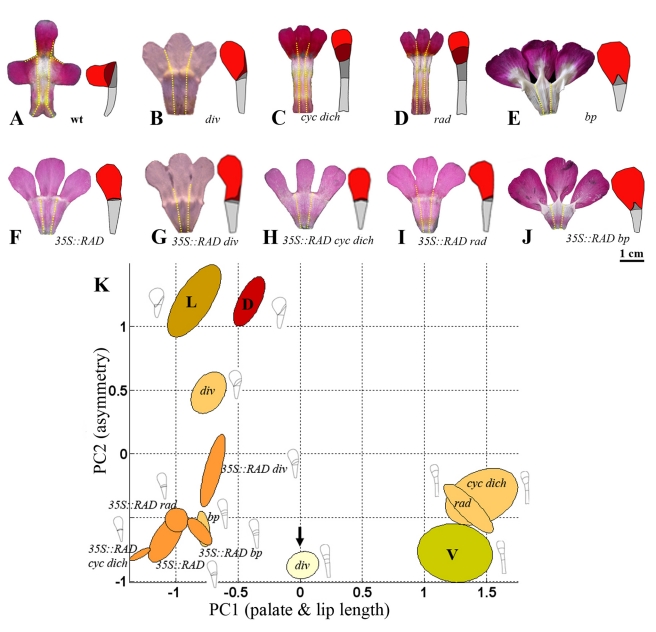
The development of organs with particular shapes, like wings or flowers, depends on regional activity of transcription factors and signalling molecules. However, the mechanisms that link these molecular activities to the morphogenetic events underlying shape are poorly understood. Here we describe a combination of experimental and computational approaches that address this problem, applying them to a group of genes controlling flower shape in the Snapdragon (Antirrhinum). Four transcription factors are known to play a key role in the control of floral shape and asymmetry in Snapdragon. We use quantitative shape analysis of mutants for these factors to define principal components underlying flower shape variation. We show that each transcription factor has a specific effect on the shape and size of regions within the flower, shifting the position of the flower in shape space. These shifts are further analysed by generating double mutants and lines that express some of the genes ectopically. By integrating these observations with known gene expression patterns and interactions, we arrive at a combinatorial scheme for how regional effects on shape are genetically controlled. We evaluate our scheme by incorporating the proposed interactions into a generative model, where the developing flower is treated as a material sheet that grows according to how genes modify local polarities and growth rates. The petal shapes generated by the model show a good quantitative match with those observed experimentally for each petal in numerous genotypes, thus validating the hypothesised scheme. This article therefore shows how complex shapes can be accounted for by combinatorial effects of transcription factors on regional growth properties. This finding has implications not only for how shapes develop but also for how they may have evolved through tinkering with transcription factors and their targets.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures







References
-
- Held L. I. UK: Cambridge University Press; 2002. Imaginal discs: the genetic and cellular logic of pattern formation:
-
- Hejnowicz Z, Romberger J. A. Growth tensor of plant organs. Journal of Theoretical Biology. 1984;110:93–114.
-
- Sinnott E. New York: McGraw-Hill; 1960. Plant morphogenesis:
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
