

Differential effects of injury severity on cognition and cellular pathology after contusive brain trauma in the immature rat
- PMID: 21091272
- PMCID: PMC3037812
- DOI: 10.1089/neu.2010.1639
Differential effects of injury severity on cognition and cellular pathology after contusive brain trauma in the immature rat
Abstract
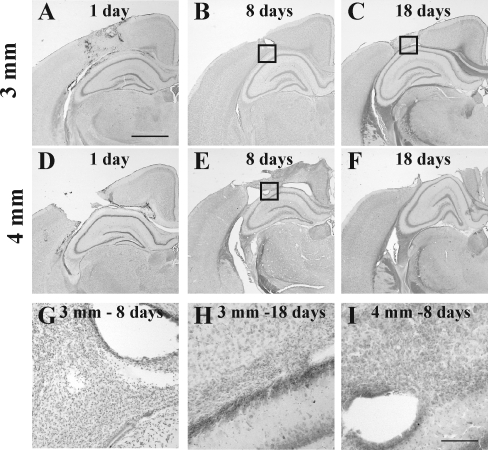
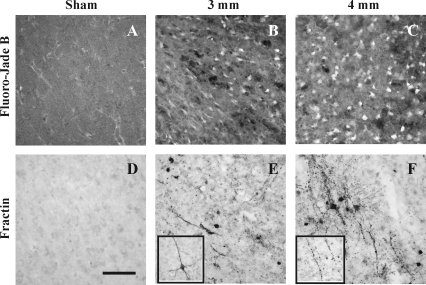
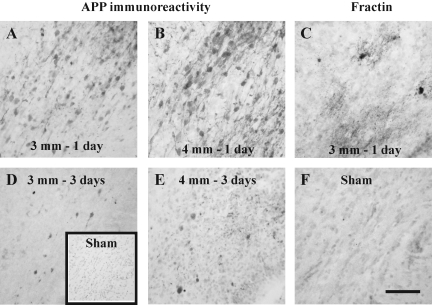
Although diffuse brain damage has been suggested to be the predominant predictor of neurological morbidity following closed head injury in infants and children, the presence of contusions also predicts long-term neurobehavioral dysfunction. Contusive brain trauma in the 17-day-old rat resulted in neurodegeneration and caspase activation in the cortex at 1 day, and in the thalamus at 3 days post-injury, and to a greater extent following a deeper impact. Cortical tissue loss in the 4-mm impact group was significantly greater than that in the 3-mm impact group (p < 0.05), and exhibited a time-dependent increase over the first 3 weeks post-injury. Traumatic axonal injury was observed in the white matter tracts below the site of impact at 1 day, and in the corpus callosum at 3 days, to a greater extent following 4-mm impact. In contrast, cellular caspase-3 activation in these white matter tracts was only observed at 24 h post-injury and was not affected by impact depth. Similarly, neurodegeneration and caspase activation in the hippocampus was restricted to the dentate gyrus and occurred to a similar extent in both injured groups. Only the 4-mm impact group exhibited learning deficits in the first week (p < 0.0001) that was sustained until the third week post-injury (p < 0.0001), while deficits in the 3-mm impact group were seen only at 3 weeks post-injury (p < 0.02). These observations demonstrate that increasing severity of injury in immature animals does not uniformly increase the extent of cellular damage, and that the progression of tissue damage and behavioral deficits varies as a function of injury severity.
Figures





References
-
- Adelson P.D. Dixon C.E. Robichaud P. Kochanek P.M. Motor and cognitive functional deficits following diffuse traumatic brain injury in the immature rat. J. Neurotrauma. 1997;14:99–108. - PubMed
-
- Adelson P.D. Whalen M.J. Kochanek P.M. Robichaud P. Carlos T.M. Blood-brain barrier permeability and acute inflammation in two models of traumatic brain injury in the immature rat: a preliminary report. Acta Neurochir. Suppl. 1998;71:104–106. - PubMed
-
- Anderson V.A. Catroppa C. Rosenfeld J. Haritou F. Morse S.A. Recovery of memory function following traumatic brain injury in pre-school children. Brain Inj. 2000;14:679–692. - PubMed
-
- Anderson V. Catroppa C. Morse S. Haritou F. Rosenfeld J. Functional plasticity or vulnerability after early brain injury? Pediatrics. 2005;116:1374–1382. - PubMed
-
- Arbogast K.B. Margulies S.S. Christian C.W. Initial neurologic presentation in young children sustaining inflicted and unintentional fatal head injuries. Pediatrics. 2005;116:180–184. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
