

Carbon flux rerouting during Mycobacterium tuberculosis growth arrest
- PMID: 21091505
- PMCID: PMC3072047
- DOI: 10.1111/j.1365-2958.2010.07399.x
Carbon flux rerouting during Mycobacterium tuberculosis growth arrest
Erratum in
-
Carbon flux rerouting during Mycobacterium tuberculosis growth arrest.Mol Microbiol. 2016 Mar;99(6):1179. doi: 10.1111/mmi.13350. Mol Microbiol. 2016. PMID: 26971532 No abstract available.
Abstract
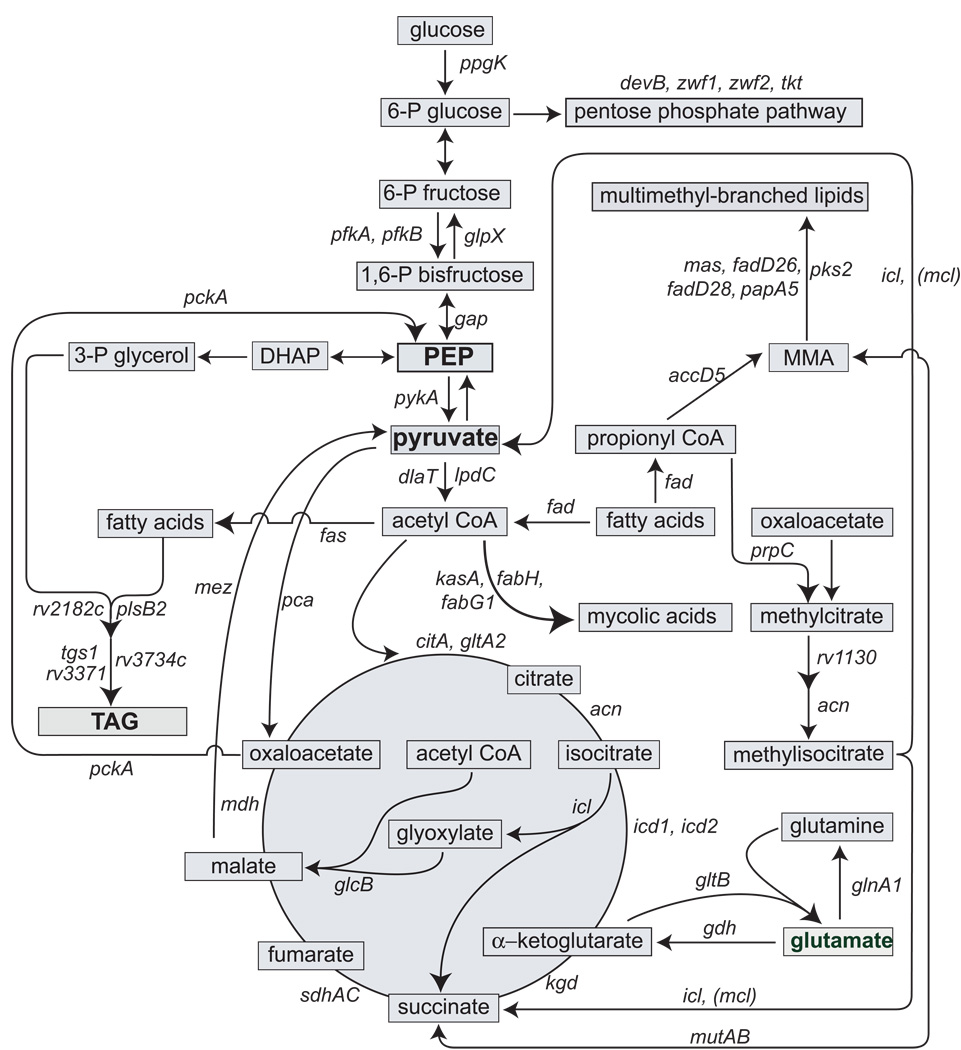
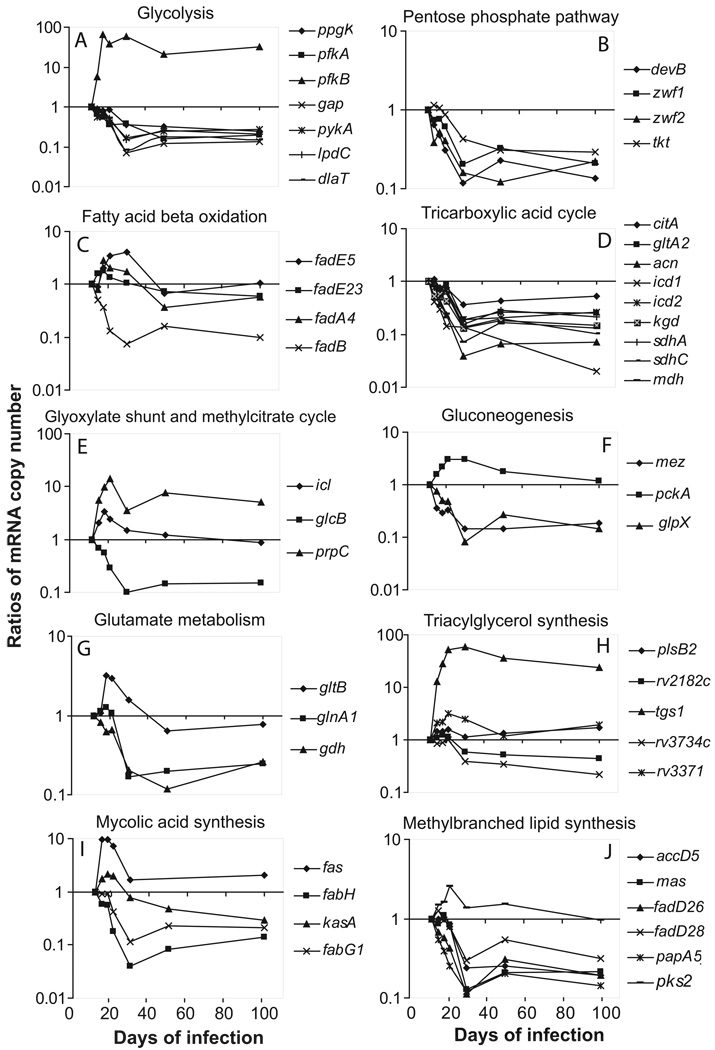
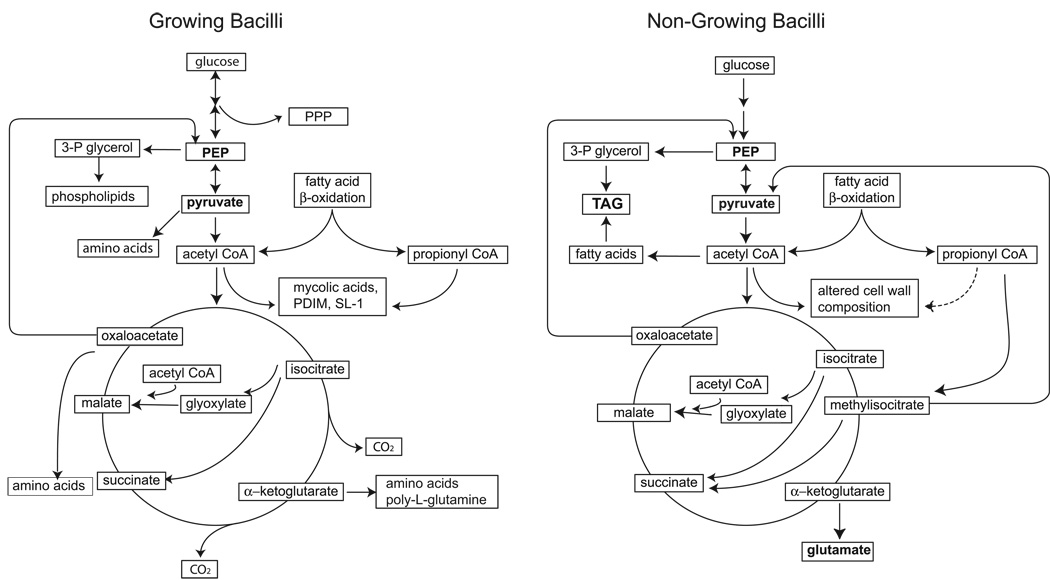
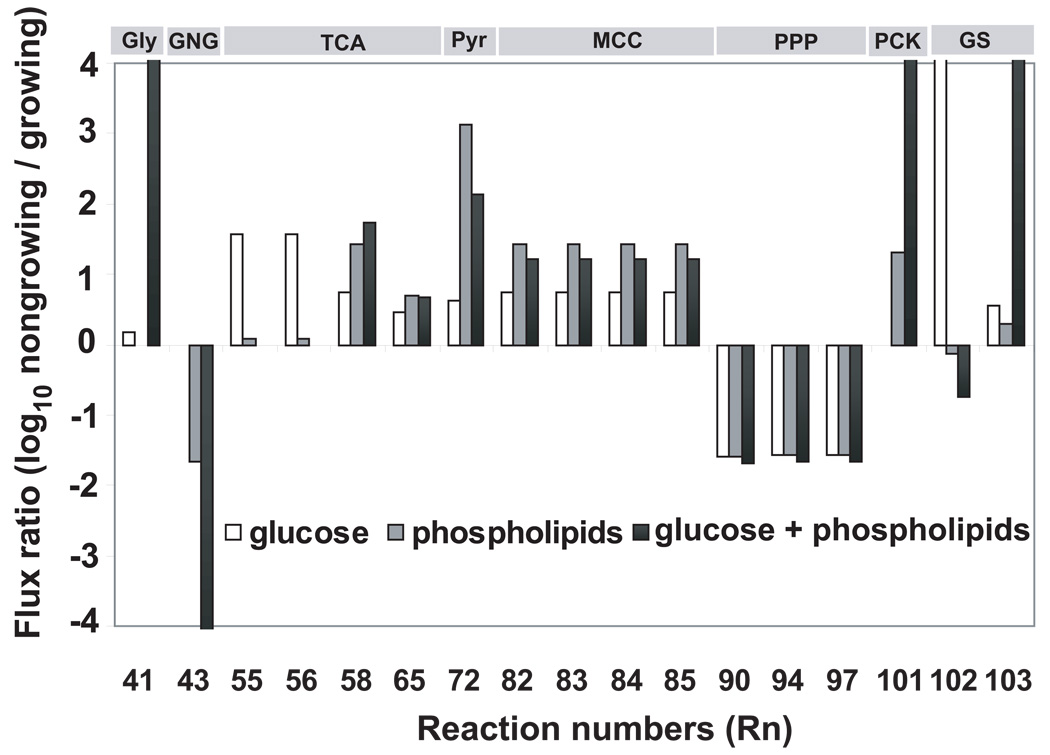
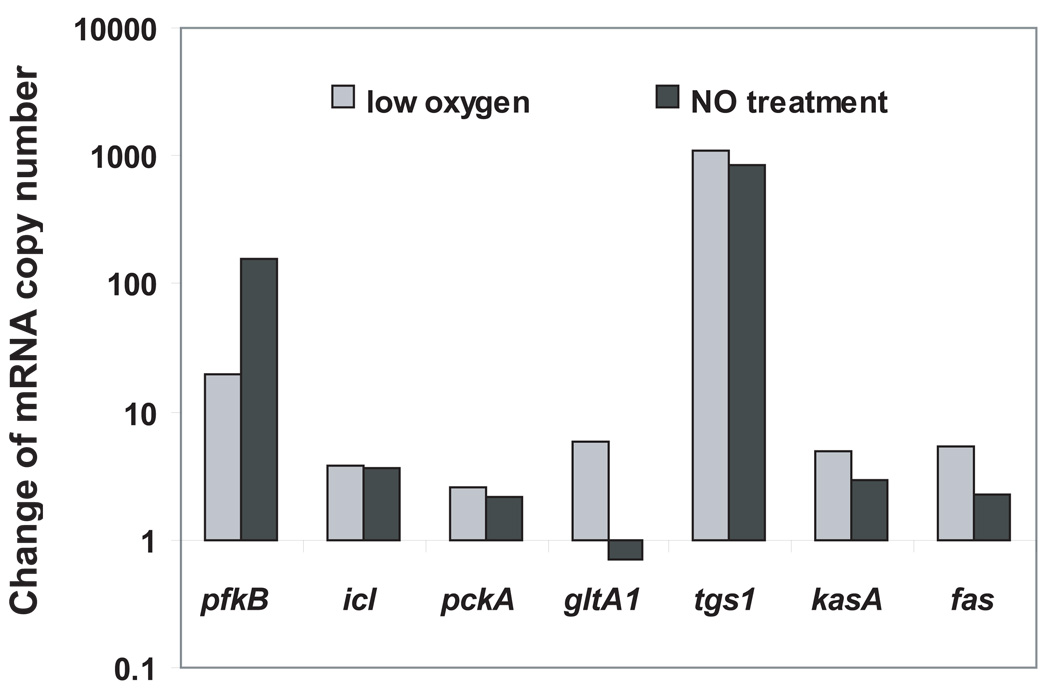
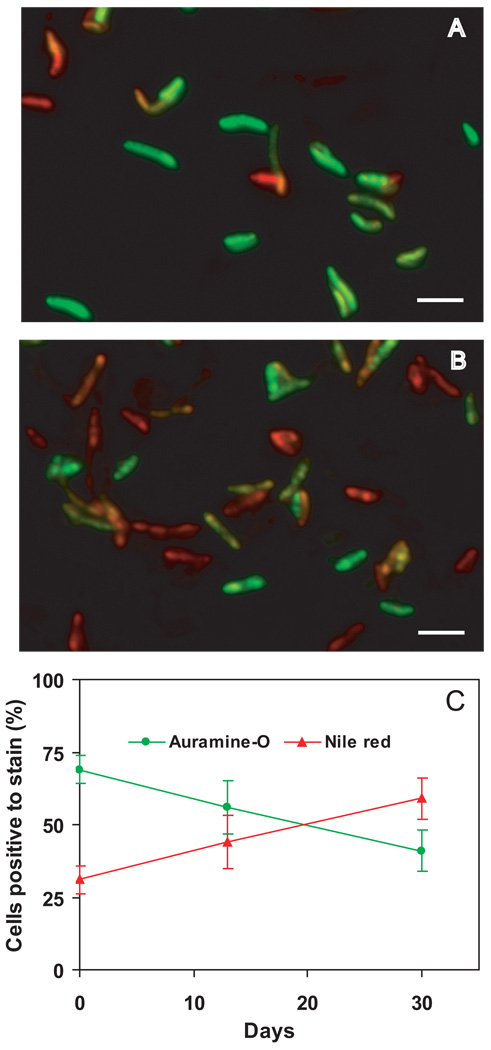
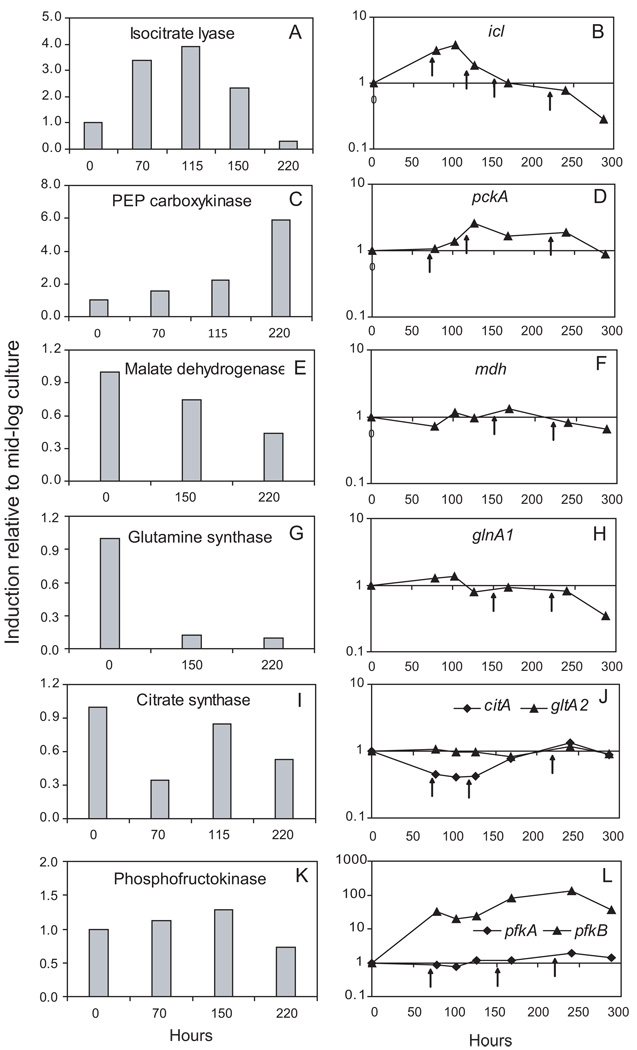
A hallmark of the Mycobacterium tuberculosis life cycle is the pathogen's ability to switch between replicative and non-replicative states in response to host immunity. Transcriptional profiling by qPCR of ∼ 50 M. tuberculosis genes involved in central and lipid metabolism revealed a re-routing of carbon flow associated with bacterial growth arrest during mouse lung infection. Carbon rerouting was marked by a switch from metabolic pathways generating energy and biosynthetic precursors in growing bacilli to pathways for storage compound synthesis during growth arrest. Results of flux balance analysis using an in silico metabolic network were consistent with the transcript abundance data obtained in vivo. Similar transcriptional changes were seen in vitro when M. tuberculosis cultures were treated with bacteriostatic stressors under different nutritional conditions. Thus, altered expression of key metabolic genes reflects growth rate changes rather than changes in substrate availability. A model describing carbon flux rerouting was formulated that (i) provides a coherent interpretation of the adaptation of M. tuberculosis metabolism to immunity-induced stress and (ii) identifies features common to mycobacterial dormancy and stress responses of other organisms.
© 2010 Blackwell Publishing Ltd.
Figures
Similar articles
-
Role of Premycofactocin Synthase in Growth, Microaerophilic Adaptation, and Metabolism of Mycobacterium tuberculosis.mBio. 2021 Aug 31;12(4):e0166521. doi: 10.1128/mBio.01665-21. Epub 2021 Jul 27. mBio. 2021. PMID: 34311585 Free PMC article.
-
Slow growth of Mycobacterium tuberculosis at acidic pH is regulated by phoPR and host-associated carbon sources.Mol Microbiol. 2014 Oct;94(1):56-69. doi: 10.1111/mmi.12688. Epub 2014 Jul 13. Mol Microbiol. 2014. PMID: 24975990 Free PMC article.
-
Comparative proteomic profiles reveal characteristic Mycobacterium tuberculosis proteins induced by cholesterol during dormancy conditions.Microbiology (Reading). 2017 Aug;163(8):1237-1247. doi: 10.1099/mic.0.000512. Epub 2017 Aug 4. Microbiology (Reading). 2017. PMID: 28771131
-
Triacylglycerol: nourishing molecule in endurance of Mycobacterium tuberculosis.J Biosci. 2018 Mar;43(1):149-154. J Biosci. 2018. PMID: 29485123 Review.
-
Mycobacterial genes essential for the pathogen's survival in the host.Immunol Rev. 2015 Mar;264(1):319-26. doi: 10.1111/imr.12256. Immunol Rev. 2015. PMID: 25703569 Free PMC article. Review.
Cited by
-
Structural and functional studies of phosphoenolpyruvate carboxykinase from Mycobacterium tuberculosis.PLoS One. 2015 Mar 23;10(3):e0120682. doi: 10.1371/journal.pone.0120682. eCollection 2015. PLoS One. 2015. PMID: 25798914 Free PMC article.
-
Triacylglycerols: Fuelling the Hibernating Mycobacterium tuberculosis.Front Cell Infect Microbiol. 2019 Jan 9;8:450. doi: 10.3389/fcimb.2018.00450. eCollection 2018. Front Cell Infect Microbiol. 2019. PMID: 30687647 Free PMC article. Review.
-
Biphasic Dynamics of Macrophage Immunometabolism during Mycobacterium tuberculosis Infection.mBio. 2019 Mar 26;10(2):e02550-18. doi: 10.1128/mBio.02550-18. mBio. 2019. PMID: 30914513 Free PMC article. Review.
-
Clonal Diversification and Changes in Lipid Traits and Colony Morphology in Mycobacterium abscessus Clinical Isolates.J Clin Microbiol. 2015 Nov;53(11):3438-47. doi: 10.1128/JCM.02015-15. Epub 2015 Aug 19. J Clin Microbiol. 2015. PMID: 26292297 Free PMC article.
-
Biochemical analysis of the NAD+-dependent malate dehydrogenase, a substrate of several serine/threonine protein kinases of Mycobacterium tuberculosis.PLoS One. 2015 Apr 10;10(4):e0123327. doi: 10.1371/journal.pone.0123327. eCollection 2015. PLoS One. 2015. PMID: 25860441 Free PMC article.
References
-
- Alvarez HM, Steinbuchel A. Triacylglycerols in prokaryotic microorganisms. Applied microbiology and biotechnology. 2002;60:367–376. - PubMed
-
- Bhatt A, Molle V, Besra GS, Jacobs WR, Jr, Kremer L. The Mycobacterium tuberculosis FAS-II condensing enzymes: their role in mycolic acid biosynthesis, acid-fastness, pathogenesis and in future drug development. Molecular microbiology. 2007;64:1442–1454. - PubMed
-
- Bishai W. Lipid lunch for persistent pathogen. Nature. 2000;406:683–685. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
