

Translational independence between overlapping genes for a restriction endonuclease and its transcriptional regulator
- PMID: 21092102
- PMCID: PMC2997769
- DOI: 10.1186/1471-2199-11-87
Translational independence between overlapping genes for a restriction endonuclease and its transcriptional regulator
Abstract
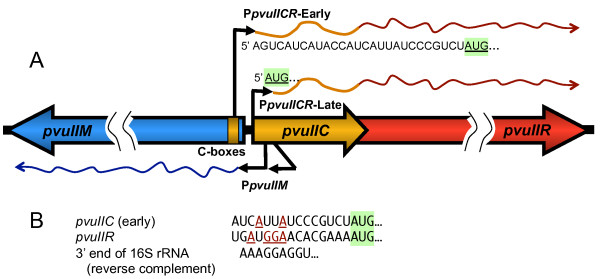
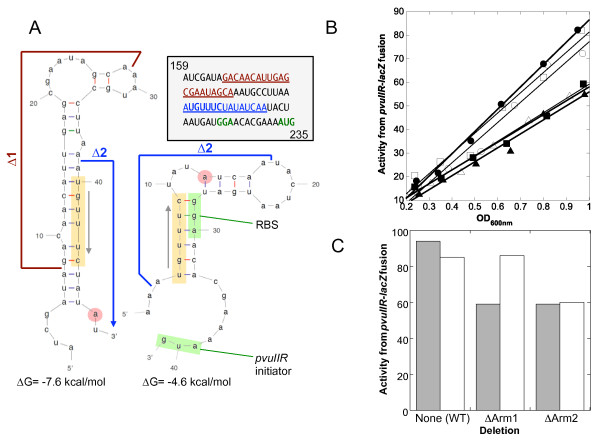
Background: Most type II restriction-modification (RM) systems have two independent enzymes that act on the same DNA sequence: a modification methyltransferase that protects target sites, and a restriction endonuclease that cleaves unmethylated target sites. When RM genes enter a new cell, methylation must occur before restriction activity appears, or the host's chromosome is digested. Transcriptional mechanisms that delay endonuclease expression have been identified in some RM systems. A substantial subset of those systems is controlled by a family of small transcription activators called C proteins. In the PvuII system, C.PvuII activates transcription of its own gene, along with that of the downstream endonuclease gene. This regulation results in very low R.PvuII mRNA levels early after gene entry, followed by rapid increase due to positive feedback. However, given the lethal consequences of premature REase accumulation, transcriptional control alone might be insufficient. In C-controlled RM systems, there is a ± 20 nt overlap between the C termination codon and the R (endonuclease) initiation codon, suggesting possible translational coupling, and in many cases predicted RNA hairpins could occlude the ribosome binding site for the endonuclease gene.
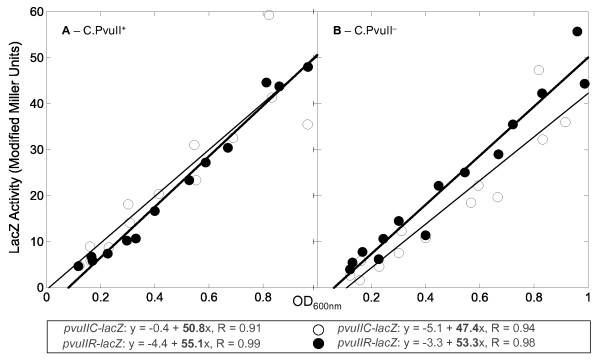
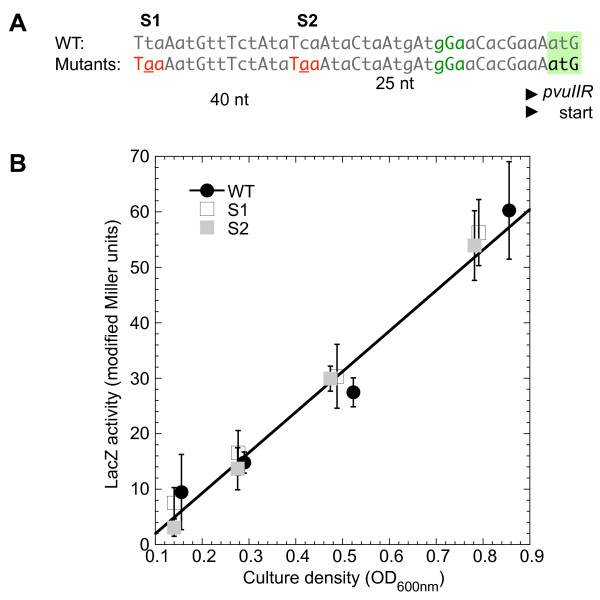
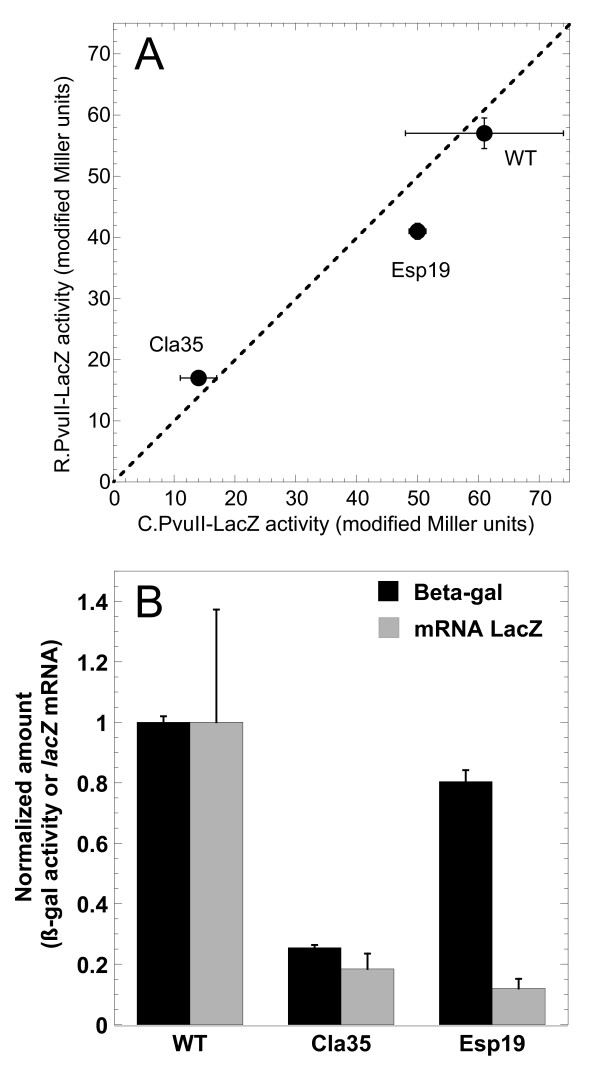
Results: Expression levels of lacZ translational fusions to pvuIIR or pvuIIC were determined, with the native pvuII promoter having been replaced by one not controlled by C.PvuII. In-frame pvuIIC insertions did not substantially decrease either pvuIIC-lacZ or pvuIIR-lacZ expression (with or without C.PvuII provided in trans). In contrast, a frameshift mutation in pvuIIC decreased expression markedly in both fusions, but mRNA measurements indicated that this decrease could be explained by transcriptional polarity. Expression of pvuIIR-lacZ was unaffected when the pvuIIC stop codon was moved 21 nt downstream from its WT location, or 25 or 40 bp upstream of the pvuIIR initiation codon. Disrupting the putative hairpins had no significant effects.
Conclusions: The initiation of translation of pvuIIR appears to be independent of that for pvuIIC. Direct tests failed to detect regulatory rules for either gene overlap or the putative hairpins. Thus, at least during balanced growth, transcriptional control appears to be sufficiently robust for proper regulation of this RM system.
Figures


Similar articles
-
Naturally-occurring, dually-functional fusions between restriction endonucleases and regulatory proteins.BMC Evol Biol. 2013 Oct 2;13:218. doi: 10.1186/1471-2148-13-218. BMC Evol Biol. 2013. PMID: 24083337 Free PMC article.
-
Role and mechanism of action of C. PvuII, a regulatory protein conserved among restriction-modification systems.J Bacteriol. 2000 Jan;182(2):477-87. doi: 10.1128/JB.182.2.477-487.2000. J Bacteriol. 2000. PMID: 10629196 Free PMC article.
-
Sequence and characterization of pvuIIR, the PvuII endonuclease gene, and of pvuIIC, its regulatory gene.J Bacteriol. 1992 May;174(10):3395-8. doi: 10.1128/jb.174.10.3395-3398.1992. J Bacteriol. 1992. PMID: 1577705 Free PMC article.
-
Nature of the promoter activated by C.PvuII, an unusual regulatory protein conserved among restriction-modification systems.J Bacteriol. 2005 Jan;187(2):488-97. doi: 10.1128/JB.187.2.488-497.2005. J Bacteriol. 2005. PMID: 15629920 Free PMC article.
-
Restriction modification systems in archaea: A panoramic outlook.Heliyon. 2024 Mar 5;10(8):e27382. doi: 10.1016/j.heliyon.2024.e27382. eCollection 2024 Apr 30. Heliyon. 2024. PMID: 38644887 Free PMC article. Review.
Cited by
-
Effect of the luxI/R gene on AHL-signaling molecules and QS regulatory mechanism in Hafnia alvei H4.AMB Express. 2019 Dec 5;9(1):197. doi: 10.1186/s13568-019-0917-z. AMB Express. 2019. PMID: 31807954 Free PMC article.
-
Natural C-independent expression of restriction endonuclease in a C protein-associated restriction-modification system.Nucleic Acids Res. 2016 Apr 7;44(6):2646-60. doi: 10.1093/nar/gkv1331. Epub 2015 Dec 9. Nucleic Acids Res. 2016. PMID: 26656489 Free PMC article.
-
Naturally-occurring, dually-functional fusions between restriction endonucleases and regulatory proteins.BMC Evol Biol. 2013 Oct 2;13:218. doi: 10.1186/1471-2148-13-218. BMC Evol Biol. 2013. PMID: 24083337 Free PMC article.
-
Overlapping genes: a window on gene evolvability.BMC Genomics. 2014 Aug 27;15(1):721. doi: 10.1186/1471-2164-15-721. BMC Genomics. 2014. PMID: 25159814 Free PMC article.
-
Chromosomal Arrangement of AHL-Driven Quorum Sensing Circuits in Pseudomonas.ISRN Microbiol. 2012 Feb 29;2012:484176. doi: 10.5402/2012/484176. Print 2012. ISRN Microbiol. 2012. PMID: 23724324 Free PMC article.
References
-
- Releigh EA, Brooks JE. Restriction Modification systems: where they are and what they do. Chapman & Hall, New York, N.Y; 1998.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
