

Combining nitric oxide release with anti-inflammatory activity preserves nigrostriatal dopaminergic innervation and prevents motor impairment in a 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine model of Parkinson's disease
- PMID: 21092260
- PMCID: PMC3000390
- DOI: 10.1186/1742-2094-7-83
Combining nitric oxide release with anti-inflammatory activity preserves nigrostriatal dopaminergic innervation and prevents motor impairment in a 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine model of Parkinson's disease
Abstract
Background: Current evidence suggests a role of neuroinflammation in the pathogenesis of Parkinson's disease (PD) and in the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) model of basal ganglia injury. Reportedly, nonsteroidal anti-inflammatory drugs (NSAIDs) mitigate DAergic neurotoxicity in rodent models of PD. Consistent with these findings, epidemiological analysis indicated that certain NSAIDs may prevent or delay the progression of PD. However, a serious impediment of chronic NSAID therapy, particularly in the elderly, is gastric, renal and cardiac toxicity. Nitric oxide (NO)-donating NSAIDs, have a safer profile while maintaining anti-inflammatory activity of parent compounds. We have investigated the oral activity of the NO-donating derivative of flurbiprofen, [2-fluoro-α-methyl (1,1'-biphenyl)-4-acetic-4-(nitrooxy)butyl ester], HCT1026 (30 mg kg(-1) daily in rodent chow) in mice exposed to the parkinsonian neurotoxin MPTP.
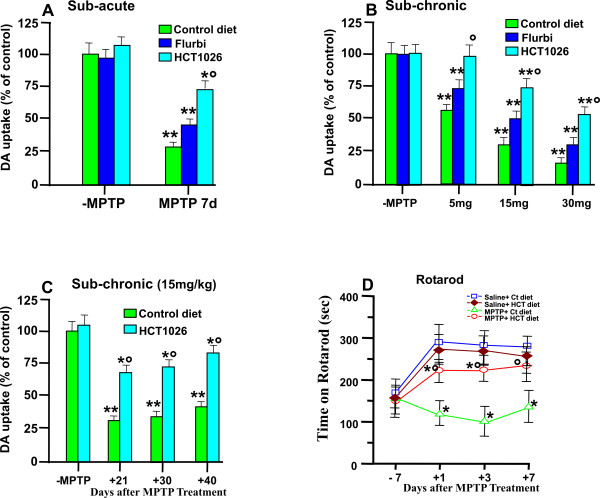
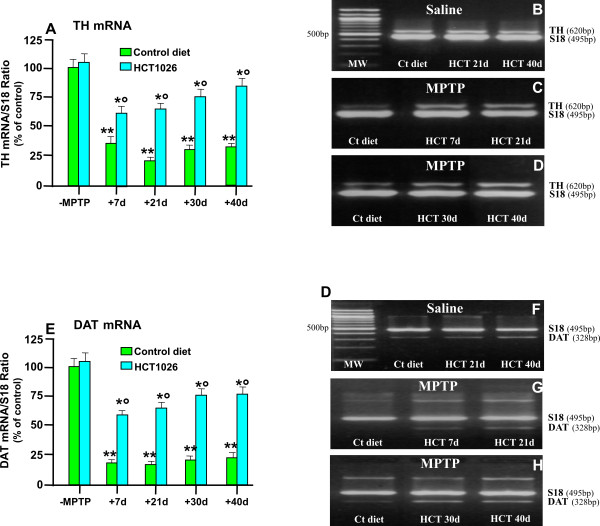
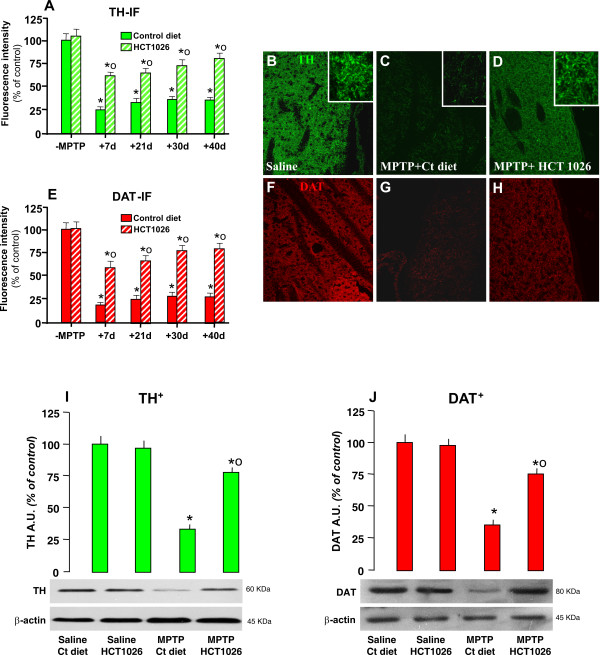
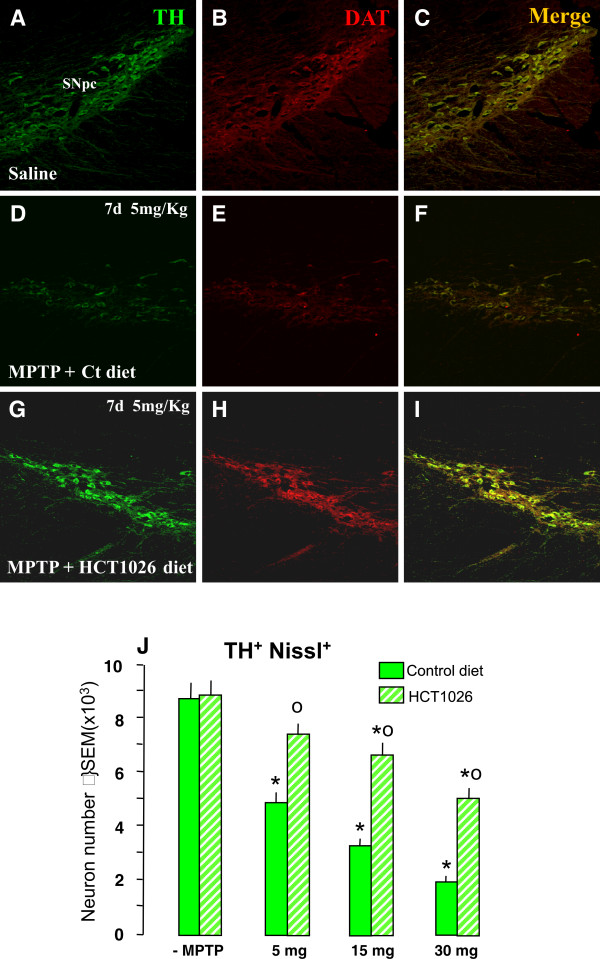
Methods: Ageing mice were fed with a control, flurbiprofen, or HCT1026 diet starting ten days before MPTP administration and continuing for all the experimental period. Striatal high affinity synaptosomal dopamine up-take, motor coordination assessed with the rotarod, tyrosine hydroxylase (TH)- and dopamine transporter (DAT) fiber staining, stereological cell counts, immunoblotting and gene expression analyses were used to assess MPTP-induced nigrostriatal DAergic toxicity and glial activation 1-40 days post-MPTP.
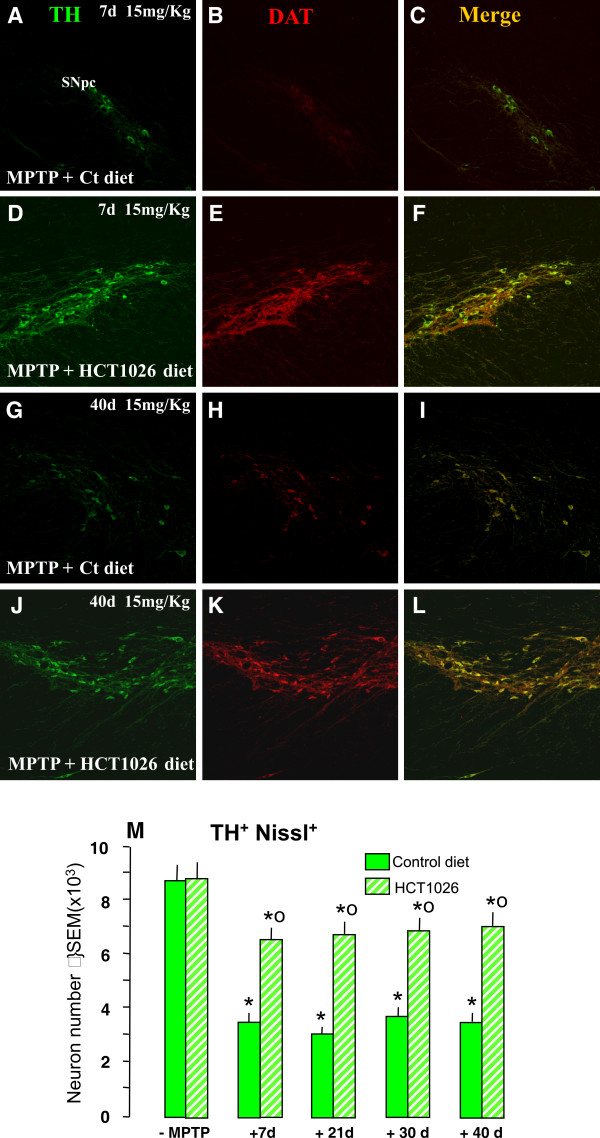
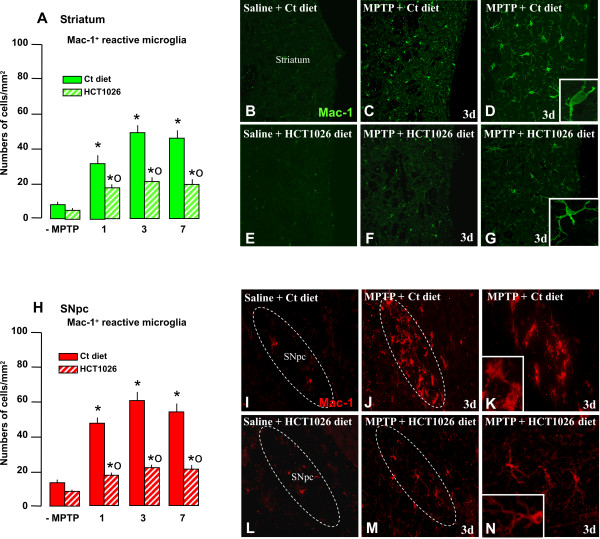
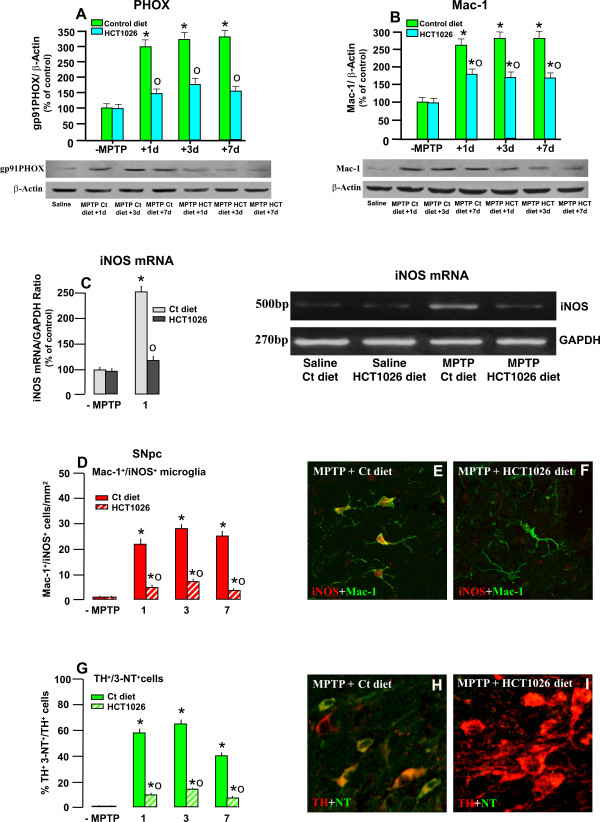
Results: HCT1026 was well tolerated and did not cause any measurable toxic effect, whereas flurbiprofen fed mice showed severe gastrointestinal side-effects. HCT1026 efficiently counteracted motor impairment and reversed MPTP-induced decreased synaptosomal [3H]dopamine uptake, TH- and DAT-stained fibers in striatum and TH+ neuron loss in substantia nigra pars compacta (SNpc), as opposed to age-matched mice fed with a control diet. These effects were associated to a significant decrease in reactive macrophage antigen-1 (Mac-1)-positive microglial cells within the striatum and ventral midbrain, decreased expression of iNOS, Mac-1 and NADPH oxidase (PHOX), and downregulation of 3-Nitrotyrosine, a peroxynitrite finger print, in SNpc DAergic neurons.
Conclusions: Oral treatment with HCT1026 has a safe profile and a significant efficacy in counteracting MPTP-induced dopaminergic (DAergic) neurotoxicity, motor impairment and microglia activation in ageing mice. HCT1026 provides a novel promising approach towards the development of effective pharmacological neuroprotective strategies against PD.
Figures
Similar articles
-
Social enrichment attenuates nigrostriatal lesioning and reverses motor impairment in a progressive 1-methyl-2-phenyl-1,2,3,6-tetrahydropyridine (MPTP) mouse model of Parkinson's disease.Neurobiol Dis. 2012 Mar;45(3):1051-67. doi: 10.1016/j.nbd.2011.12.024. Epub 2011 Dec 14. Neurobiol Dis. 2012. PMID: 22198503
-
Anti-inflammatory and neuroprotective effects of an orally active apocynin derivative in pre-clinical models of Parkinson's disease.J Neuroinflammation. 2012 Oct 23;9:241. doi: 10.1186/1742-2094-9-241. J Neuroinflammation. 2012. PMID: 23092448 Free PMC article.
-
Neuroprotective effect of ghrelin in the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine mouse model of Parkinson's disease by blocking microglial activation.Neurotox Res. 2009 May;15(4):332-47. doi: 10.1007/s12640-009-9037-x. Epub 2009 Mar 17. Neurotox Res. 2009. PMID: 19384567
-
The intranasal administration of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP): a new rodent model to test palliative and neuroprotective agents for Parkinson's disease.Curr Pharm Des. 2011;17(5):489-507. doi: 10.2174/138161211795164095. Curr Pharm Des. 2011. PMID: 21375482 Review.
-
Calpain activation and progression of inflammatory cycles in Parkinson's disease.Front Biosci (Landmark Ed). 2022 Jan 13;27(1):20. doi: 10.31083/j.fbl2701020. Front Biosci (Landmark Ed). 2022. PMID: 35090325 Free PMC article. Review.
Cited by
-
Glia-Derived Extracellular Vesicles in Parkinson's Disease.J Clin Med. 2020 Jun 21;9(6):1941. doi: 10.3390/jcm9061941. J Clin Med. 2020. PMID: 32575923 Free PMC article. Review.
-
Proteomic Analysis of the Effect of Korean Red Ginseng in the Striatum of a Parkinson's Disease Mouse Model.PLoS One. 2016 Oct 27;11(10):e0164906. doi: 10.1371/journal.pone.0164906. eCollection 2016. PLoS One. 2016. PMID: 27788166 Free PMC article.
-
Neuroprotective effects of Bacopa monnieri in Parkinson's disease model.Metab Brain Dis. 2020 Mar;35(3):517-525. doi: 10.1007/s11011-019-00526-w. Epub 2019 Dec 13. Metab Brain Dis. 2020. PMID: 31834548
-
Stomaching the Possibility of a Pathogenic Role for Helicobacter pylori in Parkinson's Disease.J Parkinsons Dis. 2018;8(3):367-374. doi: 10.3233/JPD-181327. J Parkinsons Dis. 2018. PMID: 29966206 Free PMC article. Review.
-
Targeting Wnt signaling at the neuroimmune interface for dopaminergic neuroprotection/repair in Parkinson's disease.J Mol Cell Biol. 2014 Feb;6(1):13-26. doi: 10.1093/jmcb/mjt053. Epub 2014 Jan 14. J Mol Cell Biol. 2014. PMID: 24431301 Free PMC article. Review.
References
-
- Tedroff JM. Functional consequences of dopaminergic degeneration in Parkinson's disease. Adv Neurol. 1999;80:67–70. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
