

Creating designed zinc-finger nucleases with minimal cytotoxicity
- PMID: 21094162
- PMCID: PMC3017627
- DOI: 10.1016/j.jmb.2010.10.043
Creating designed zinc-finger nucleases with minimal cytotoxicity
Abstract
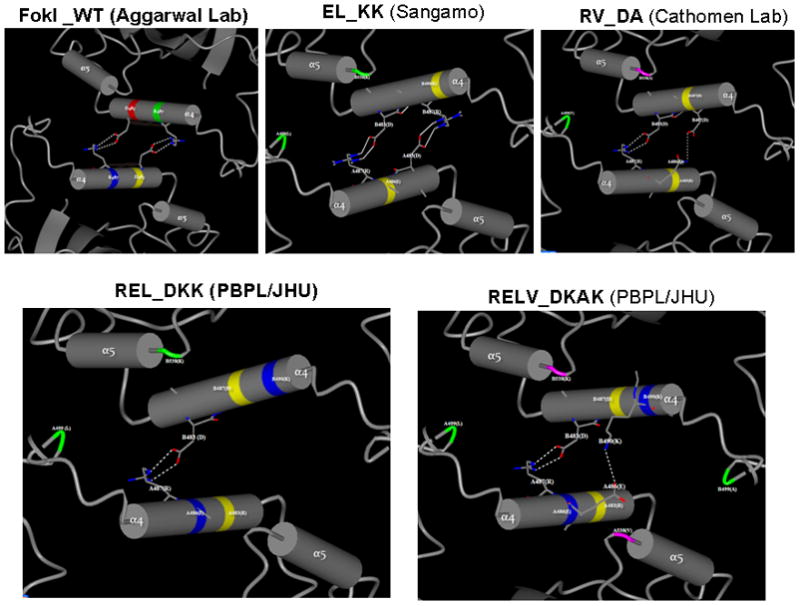
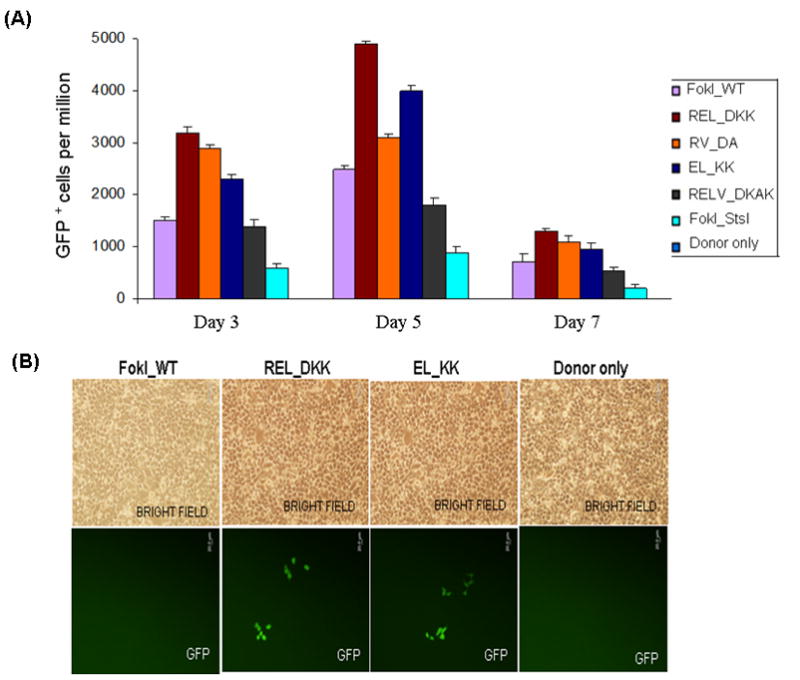
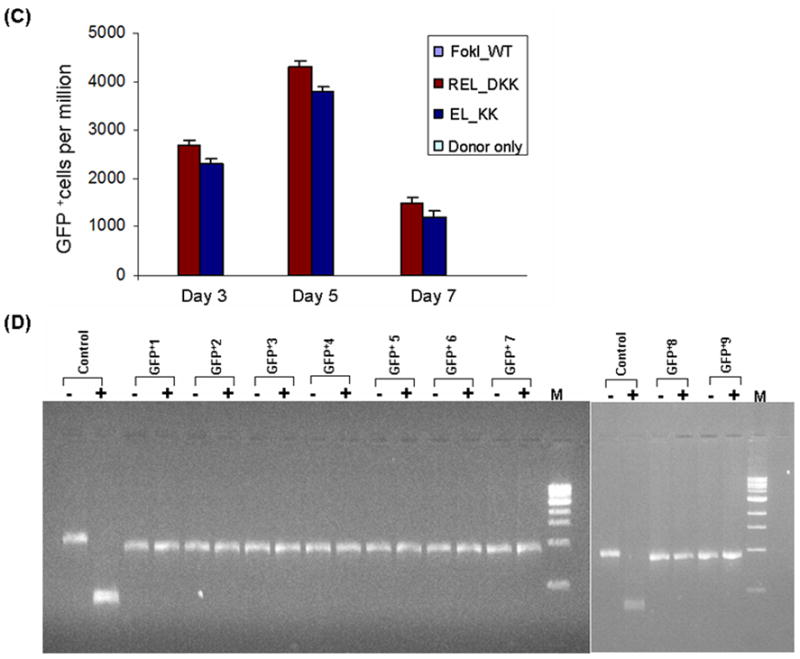
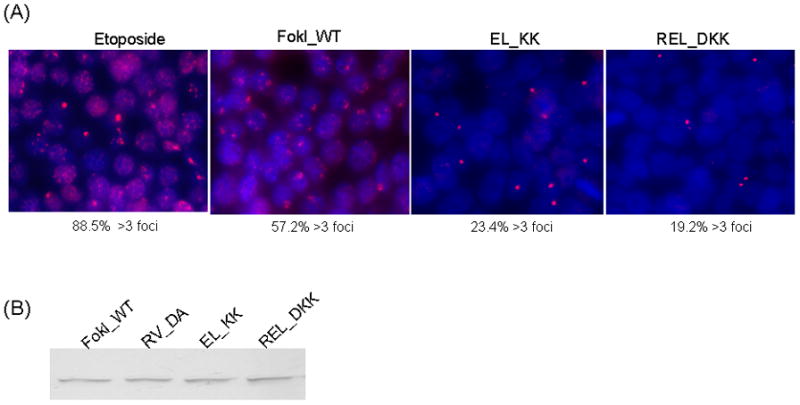
Zinc-finger nucleases (ZFNs) have emerged as powerful tools for delivering a targeted genomic double-strand break (DSB) to either stimulate local homologous recombination with investigator-provided donor DNA or induce gene mutations at the site of cleavage in the absence of a donor by nonhomologous end joining both in plant cells and in mammalian cells, including human cells. ZFNs are formed by fusing zinc-finger proteins to the nonspecific cleavage domain of the FokI restriction enzyme. ZFN-mediated gene targeting yields high gene modification efficiencies (>10%) in a variety of cells and cell types by delivering a recombinogenic DSB to the targeted chromosomal locus, using two designed ZFNs. The mechanism of DSB by ZFNs requires (1) two ZFN monomers to bind to their adjacent cognate sites on DNA and (2) the FokI nuclease domains to dimerize to form the active catalytic center for the induction of the DSB. In the case of ZFNs fused to wild-type FokI cleavage domains, homodimers may also form; this could limit the efficacy and safety of ZFNs by inducing off-target cleavage. In this article, we report further refinements to obligate heterodimer variants of the FokI cleavage domain for the creation of custom ZFNs with minimal cellular toxicity. The efficacy and efficiency of the reengineered obligate heterodimer variants of the FokI cleavage domain were tested using the green fluorescent protein gene targeting reporter system. The three-finger and four-finger zinc-finger protein fusions to the REL_DKK pair among the newly generated FokI nuclease domain variants appear to eliminate or greatly reduce the toxicity of designer ZFNs to human cells.
Copyright © 2010 Elsevier Ltd. All rights reserved.
Figures
Similar articles
-
Targeted manipulation of mammalian genomes using designed zinc finger nucleases.Biochem Biophys Res Commun. 2009 Oct 9;388(1):56-61. doi: 10.1016/j.bbrc.2009.07.112. Epub 2009 Jul 25. Biochem Biophys Res Commun. 2009. PMID: 19635463 Free PMC article.
-
Targeted gene addition to a predetermined site in the human genome using a ZFN-based nicking enzyme.Genome Res. 2012 Jul;22(7):1316-26. doi: 10.1101/gr.122879.111. Epub 2012 Mar 20. Genome Res. 2012. PMID: 22434427 Free PMC article.
-
Autonomous zinc-finger nuclease pairs for targeted chromosomal deletion.Nucleic Acids Res. 2010 Dec;38(22):8269-76. doi: 10.1093/nar/gkq720. Epub 2010 Aug 16. Nucleic Acids Res. 2010. PMID: 20716517 Free PMC article.
-
Zinc finger nucleases: custom-designed molecular scissors for genome engineering of plant and mammalian cells.Nucleic Acids Res. 2005 Oct 26;33(18):5978-90. doi: 10.1093/nar/gki912. Print 2005. Nucleic Acids Res. 2005. PMID: 16251401 Free PMC article. Review.
-
Origins of Programmable Nucleases for Genome Engineering.J Mol Biol. 2016 Feb 27;428(5 Pt B):963-89. doi: 10.1016/j.jmb.2015.10.014. Epub 2015 Oct 23. J Mol Biol. 2016. PMID: 26506267 Free PMC article. Review.
Cited by
-
Site- and strand-specific nicking of DNA by fusion proteins derived from MutH and I-SceI or TALE repeats.Nucleic Acids Res. 2013 Apr;41(7):e83. doi: 10.1093/nar/gkt080. Epub 2013 Feb 13. Nucleic Acids Res. 2013. PMID: 23408850 Free PMC article.
-
Current translational and clinical practices in hematopoietic cell and gene therapy.Cytotherapy. 2012 Aug;14(7):775-90. doi: 10.3109/14653249.2012.694420. Cytotherapy. 2012. PMID: 22799276 Free PMC article. Review.
-
Transcriptional Activation of Biosynthetic Gene Clusters in Filamentous Fungi.Front Bioeng Biotechnol. 2022 Jul 15;10:901037. doi: 10.3389/fbioe.2022.901037. eCollection 2022. Front Bioeng Biotechnol. 2022. PMID: 35910033 Free PMC article. Review.
-
Comparing zinc finger nucleases and transcription activator-like effector nucleases for gene targeting in Drosophila.G3 (Bethesda). 2013 Oct 3;3(10):1717-25. doi: 10.1534/g3.113.007260. G3 (Bethesda). 2013. PMID: 23979928 Free PMC article.
-
Obligate ligation-gated recombination (ObLiGaRe): custom-designed nuclease-mediated targeted integration through nonhomologous end joining.Genome Res. 2013 Mar;23(3):539-46. doi: 10.1101/gr.145441.112. Epub 2012 Nov 14. Genome Res. 2013. PMID: 23152450 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
