

Phosphatidylserine reduces immune response against human recombinant Factor VIII in Hemophilia A mice by regulation of dendritic cell function
- PMID: 21094627
- PMCID: PMC3059555
- DOI: 10.1016/j.clim.2010.10.006
Phosphatidylserine reduces immune response against human recombinant Factor VIII in Hemophilia A mice by regulation of dendritic cell function
Abstract
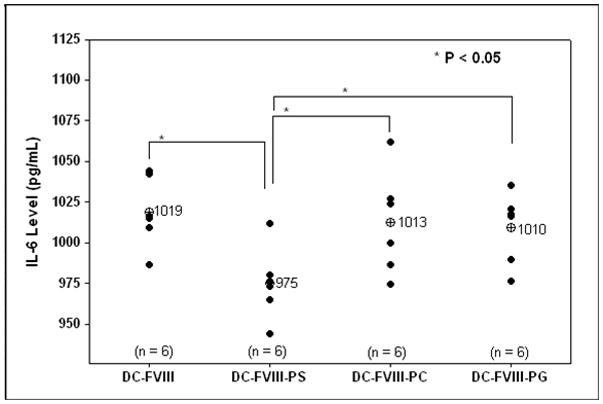
A major clinical complication in the treatment of Hemophilia A using exogenously administered recombinant Factor VIII (FVIII) is the development of neutralizing antibodies. It has been shown previously that FVIII complexed with phosphatidylserine (PS) reduces the development of total and neutralizing antibody titers in hemophilic mice. The effect of complexation of FVIII with PS upon dendritic cell (DC) uptake, maturation and processing, T-cell proliferation and cytokine secretion profiles was investigated. Flow cytometric studies of DC showed that PS inhibited the up-regulation of cell surface co-stimulatory markers (CD86 and CD40). PS reduced T-cell proliferation and significantly increased levels of TGF-β and IL-10 but reduced secretion of IL-6 and IL-17 compared to controls. The data suggest that PS reduces immunogenicity of FVIII by regulating dendritic cell maturation and subsequent T-lymphocyte activity through modulation of cytokine secretion. A possible mechanism for PS-mediated induction of FVIII tolerance is discussed.
Copyright © 2010 Elsevier Inc. All rights reserved.
Figures















References
-
- Kaufman RJ, Wasley LC, Dorner AJ. Synthesis, processing, and secretion of recombinant human factor VIII expressed in mammalian cells. J Biol Chem. 1988;263(13):6352–62. - PubMed
-
- Vehar GA, et al. Structure of human factor VIII. Nature. 1984;312(5992):337–42. - PubMed
-
- Fay PJ. Factor VIII structure and function. Thromb Haemost. 1993;70(1):63–7. - PubMed
-
- Lollar P, et al. Factor VIII inhibitors. Adv Exp Med Biol. 2001;489:65–73. - PubMed
-
- Ananyeva NM, et al. Inhibitors in hemophilia A: mechanisms of inhibition, management and perspectives. Blood Coagul Fibrinolysis. 2004;15(2):109–24. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
